Modeling Alzheimer's Disease Mechanisms with iPSCs: From Pathogenesis to Drug Discovery
This article provides a comprehensive analysis of how induced pluripotent stem cell (iPSC) models are revolutionizing Alzheimer's disease (AD) research.
Modeling Alzheimer's Disease Mechanisms with iPSCs: From Pathogenesis to Drug Discovery
Abstract
This article provides a comprehensive analysis of how induced pluripotent stem cell (iPSC) models are revolutionizing Alzheimer's disease (AD) research. It explores the foundational mechanisms of AD, including Aβ and tau pathology, as revealed by patient-specific neurons. The content details methodological advances in creating high-fidelity cortical and motor neuron models, 3D systems, and their application in high-throughput drug screening. It addresses key challenges in model standardization and reproducibility while evaluating the validation of these models against traditional systems and their predictive power for clinical outcomes. Finally, it synthesizes how iPSC technology is paving the way for personalized medicine and identifying novel therapeutic combinations for this complex neurodegenerative disorder.
Decoding Alzheimer's Pathology: How iPSC Models Illuminate Aβ, Tau, and Neurodegenerative Mechanisms
Alzheimer's disease (AD), the most common cause of dementia, presents a profound global health challenge with an estimated 7.1 million affected Americans and projections exceeding 13.9 million by 2060 [1]. For decades, Alzheimer's research has relied heavily on animal models, which often fail to fully recapitulate human-specific disease processes, contributing to the repeated failure of clinical trials [2] [3]. The discovery of induced pluripotent stem cell (iPSC) technology has revolutionized this landscape, enabling researchers to generate patient-specific human neurons and glial cells that recapitulate the complex pathophysiology of AD in a dish [4] [2].
The core pathological hallmarks of AD—extracellular amyloid plaques composed primarily of amyloid-β (Aβ) peptides and intracellular neurofibrillary tangles (NFTs) consisting of hyperphosphorylated tau protein—have been exceptionally difficult to model in conventional systems [5] [3]. iPSC-derived models now provide an unprecedented platform to study the formation of these lesions, their cellular interactions, and their functional consequences in human cells with relevant genetic backgrounds [2]. This technical guide explores the current state of recapitulating amyloid and tau pathology in iPSC-derived neural models, providing methodologies, analytical approaches, and applications for drug discovery.
Alzheimer's Pathophysiology and iPSC Modeling Approaches
Amyloid and Tau Pathology
In the amyloidogenic pathway, the amyloid precursor protein (APP) undergoes sequential proteolytic cleavage by β-secretase (BACE1) and γ-secretase, generating Aβ peptides of varying lengths [4]. The Aβ1-42 isoform is particularly pathogenic due to its high self-aggregation propensity and central role in forming amyloid plaques [6] [4]. Meanwhile, tau, a microtubule-associated protein, becomes hyperphosphorylated at numerous serine and threonine residues in AD, leading to its dissociation from microtubules and aggregation into paired helical filaments and eventually NFTs [6]. This hyperphosphorylation is regulated by a balance between kinases such as glycogen synthase kinase 3β (GSK3B) and phosphatases [6].
Familial vs. Sporadic AD Modeling
iPSC models have been developed for both familial AD (fAD) and sporadic AD (sAD). fAD models typically incorporate mutations in APP, PSEN1, or PSEN2 genes, which directly increase Aβ42 production or the Aβ42/Aβ40 ratio [6] [3]. In contrast, sAD models leverage cells from patients without known familial mutations, often focusing on genetic risk factors such as the APOE ε4 allele, the strongest genetic risk factor for late-onset AD [4] [3]. The ability to model both fAD and sAD using iPSC technology provides a platform for understanding shared and distinct pathomechanisms between these AD forms [6].
Beyond Neurons: The Role of Glial Cells
Recent advances have highlighted the importance of moving beyond neuron-centric models to incorporate glial cells. Microglia, the brain's resident immune cells, adopt distinct transcriptional states in AD, including a disease-associated microglia (DAM) phenotype enriched in neurodegenerative diseases [7]. Similarly, astrocytes contribute to Aβ clearance and neuroinflammation [4] [3]. The development of protocols to differentiate iPSCs into these cell types enables the creation of more complex co-culture systems that better mimic the brain environment [4] [3].
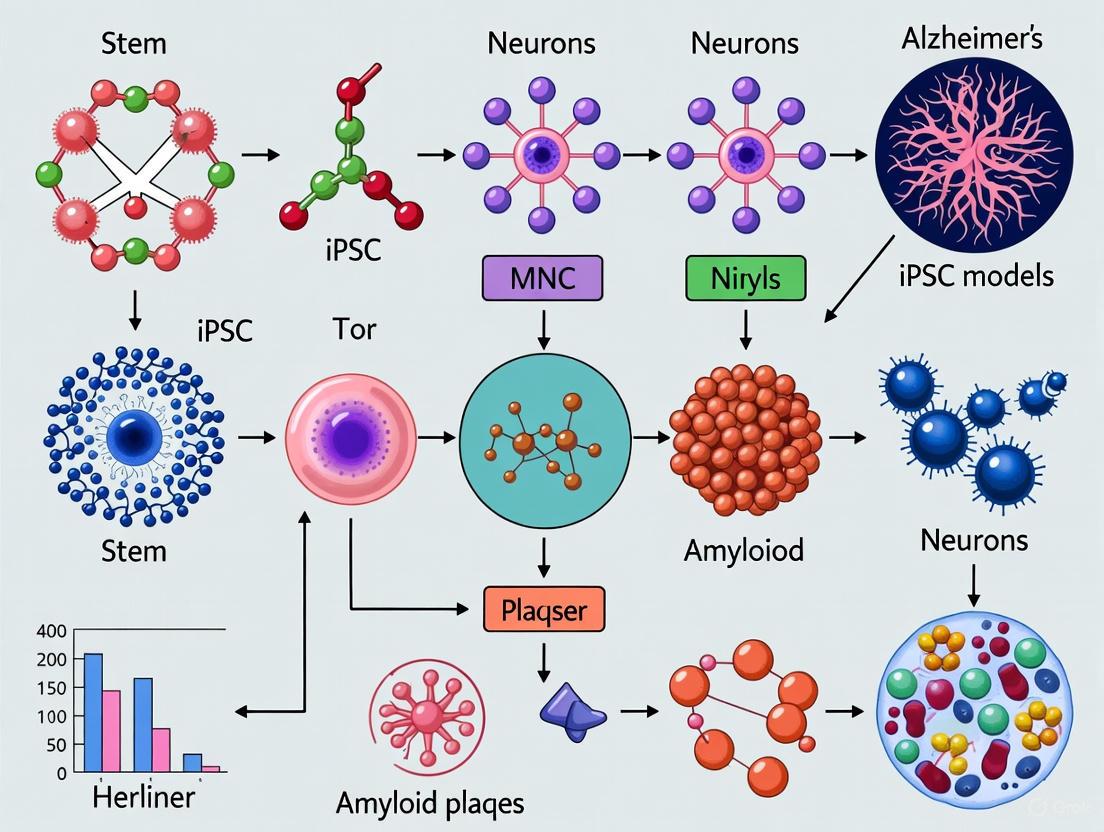
Figure 1: iPSC Modeling of Alzheimer's Disease Pathophysiology. This diagram illustrates how iPSCs can be differentiated into various neural cell types to model key AD pathologies, influenced by genetic and environmental risk factors.
Recapitulating Amyloid Pathology
Experimental Models and Methodologies
The recapitulation of amyloid pathology using iPSC-derived neurons typically involves differentiating control and AD-specific iPSCs into cortical neurons using established protocols. One common approach utilizes SMAD inhibition to direct cells toward a neural lineage, often through the formation of embryoid bodies or neural progenitor cells [2]. Neurons are then matured for several weeks to months to allow for the development of robust amyloid pathology.
For more physiologically relevant models, researchers are increasingly employing 3D culture systems such as cerebral organoids or hydrogel-based tissues that better mimic the brain's structural complexity [4] [8]. These 3D models demonstrate improved neuronal viability, network activity, and resemblance to in vivo disease pathology compared to 2D cultures [8]. A recent study described a 3D multi-electrode array (MEA) platform that enables non-invasive, real-time monitoring of extracellular field potentials throughout the entire depth of a 3D hydrogel-based neural tissue containing human iPSC-derived neurons and astrocytes [8].
Key Pathological Findings in iPSC-Derived Models
Studies using fAD iPSC-derived neurons consistently demonstrate elevated Aβ1-42 levels and increased Aβ1-42/Aβ1-40 ratios, consistent with the known effects of fAD mutations on γ-secretase processivity [6] [2]. For example, neurons with PSEN1 mutations (A246E, M146L) show significantly increased Aβ1-42/Aβ1-40 ratios compared to controls [2]. Similarly, sAD models have shown elevated levels of both extracellular Aβ1-40 and Aβ1-42, though with less consistent effects on the Aβ1-42/Aβ1-40 ratio [6].
Additionally, AD neurons exhibit increased sensitivity to oxidative stress when exposed to amyloid oligomers or peroxide, suggesting a role for amyloid in mediating oxidative damage [6]. The table below summarizes key amyloid-related phenotypes reported in various iPSC-based AD studies.
Table 1: Amyloid-β Pathology in iPSC-Derived Neuronal Models of Alzheimer's Disease
| Study Model | Aβ1-40 | Aβ1-42 | Aβ42:40 Ratio | sAPPβ | Other Phenotypes |
|---|---|---|---|---|---|
| fAD (PSEN1 A246E) [2] | ↑ | ↑ | ↑ | Increased amyloidogenic processing | |
| fAD (PSEN1 M146L) [2] | ↓ | ↑ | ↑ | - | NLRP2 inflammasome ↑ |
| fAD (APP V717L) [2] | ↑ | ↑ | ↑ | Early endosomes ↑ | |
| sAD (APOE4) [2] | ↑ | ↑ | ↑ | ↑ | Secreted APOE ↑ |
| fAD & sAD Neurons [6] | ↑ | ↑ | ↑ (fAD only) | - | GSK3B activation; Oxidative stress sensitivity ↑ |
Analysis Methods for Amyloid Pathology
- ELISA: Quantifies absolute levels of Aβ1-40, Aβ1-42, and other Aβ species in conditioned media or cell lysates [6]
- Western Blotting: Detects APP and its processing fragments, including soluble APPβ (sAPPβ) and C-terminal fragments [6]
- Immunocytochemistry: Visualizes intracellular Aβ accumulation and APP distribution using antibodies against Aβ and APP [8]
- Electrophysiology: 3D multi-electrode arrays (MEAs) monitor network-level dysfunction resulting from amyloid pathology [8]
Modeling Tau Hyperphosphorylation and Tangle Formation
Experimental Approaches to Tau Pathology
iPSC-derived neurons have successfully recapitulated the hyperphosphorylation of tau at multiple AD-relevant epitopes. A seminal study comparing fAD and sAD iPSC-derived neurons found elevated tau phosphorylation at all investigated phosphorylation sites in both AD forms, with no evident difference in phenotype between fAD and sAD samples except for secreted Aβ1-40 levels [6]. This suggests that divergent upstream triggers (e.g., increased Aβ production in fAD vs. impaired clearance in sAD) may converge on similar tau pathology mechanisms.
Notably, the GSK3B kinase has been identified as a key driver of tau hyperphosphorylation in iPSC-derived AD models. Neurons from both fAD and sAD patients show increased levels of active GSK3B, providing a mechanistic link between amyloid pathology and tau hyperphosphorylation [6]. This is further supported by observations that Aβ accumulated in the AD brain can activate kinases that promote tau phosphorylation, including GSK3B [6].
Tau Pathology in 2D vs. 3D Models
While 2D cultures have proven valuable for studying basic mechanisms of tau phosphorylation, 3D models may offer advantages for observing more advanced tau aggregation. The development of neurofibrillary tangle-like structures in iPSC-derived models has been more challenging to achieve than amyloid deposition, typically requiring extended maturation periods or additional stressors. However, increasingly complex 3D models show promise for recapitulating later stages of tau pathology [4].
Regional identity of iPSC-derived neurons also influences tau pathology, with rostral (forebrain) neuronal cells showing higher tau levels than caudal cells when derived from the same patients, highlighting the importance of neuronal subtype specification in disease modeling [2].
Table 2: Tau Pathology in iPSC-Derived Neuronal Models of Alzheimer's Disease
| Study Model | p-TAU Sites | Total TAU | GSK3B Activation | Other Phenotypes |
|---|---|---|---|---|
| fAD (PSEN1 A246E) [2] | ↑ | - | ↑ | - |
| fAD (PSEN1 V89L) [2] | ↑ | ↑ | ↑ | Sensitivity to Aβ ↑ |
| sAD (APOE4) [2] | ↑ | ↑ | - | Synapse number ↑; Early endosomes ↑ |
| fAD & sAD Neurons [6] | ↑ at multiple sites | - | ↑ | Increased sensitivity to oxidative stress |
| MAPT Mutations [2] | ↑ | - | - | ROS ↑; Firing frequency ↑ |
Analysis Methods for Tau Pathology
- Western Blotting: Using phospho-specific tau antibodies (e.g., Thr181, Ser202, Thr231) to quantify phosphorylation levels at specific epitopes [6]
- Immunocytochemistry: Visualizes spatial distribution of phosphorylated tau within neurons and its potential co-localization with other pathological markers [8]
- ELISA: Quantitative measurement of specific phosphorylated tau species in cell lysates [6]
- Synaptosome Preparation: Isolates synaptic compartments to examine synaptic tau localization and phosphorylation [7]
Advanced Model Systems: Beyond Monocultures
Co-culture Systems and Microglial Contributions
The integration of microglia into iPSC-based AD models has revealed crucial insights into neuroinflammation and plaque clearance. A groundbreaking 2023 study established a platform for generating and manipulating diverse transcriptional states in iPSC-derived human microglia (iMGLs) by exposing them to various brain substrates [7]. When iMGLs were exposed to synaptosomes, myelin debris, apoptotic neurons, or synthetic Aβ fibrils, they adopted distinct transcriptional states that mapped to signatures identified in human brain microglia, including the DAM phenotype [7].
This study further demonstrated that the transcription factor MITF drives a disease-associated transcriptional signature and a highly phagocytic state in iMGLs [7]. The ability to model these microglial states in vitro provides a powerful tool for understanding how different neural cell types interact to shape AD pathology.
3D Organoid and Tissue Engineering Approaches
3D neural tissue models show enhanced maturation of neuronal networks and more physiologically relevant cell-cell interactions compared to 2D cultures. In one advanced platform, researchers created a 3D hydrogel-based neural tissue containing human iPSC-derived neurons and primary astrocytes cultured on a custom 3D multi-electrode array [8]. This system allowed for the analysis of functional neural network development over ~6.5 weeks, revealing distinct patterns of spiking activity, bursting, and network synchrony across different depths of the 3D tissue [8].
Such 3D systems demonstrate region-specific network compositions and differential sensitivity to GABAergic and glutamatergic antagonists throughout the tissue depth, providing unprecedented resolution for studying functional network consequences of AD pathology [8].
Figure 2: Experimental Workflow for iPSC-Based AD Modeling. This diagram outlines the progressive complexity in developing iPSC-derived Alzheimer's models, from basic 2D cultures to advanced 3D systems and their subsequent applications in functional characterization and drug discovery.
The Scientist's Toolkit: Essential Research Reagents and Platforms
Table 3: Essential Research Reagents and Platforms for iPSC-Based Alzheimer's Disease Research
| Reagent/Platform | Function/Application | Examples/Specifications |
|---|---|---|
| iPSC Lines | Disease modeling with patient-specific genetics | fAD (PSEN1, APP mutations); sAD (APOE4 carriers); isogenic controls [6] [2] |
| Neural Differentiation Kits | Directing iPSC differentiation to neural lineages | SMAD inhibition protocols; Ngn2 direct induction; Embryoid body formation [2] |
| 3D Culture Matrices | Supporting three-dimensional neural tissue growth | Collagen-based hydrogels; ECM mixtures (e.g., Maxgel); Organoid culture platforms [8] |
| Aβ/Tau Assays | Quantifying amyloid and tau pathology | Aβ40/42 ELISAs; Phospho-tau Western antibodies (Thr231, Ser202); p-Tau ELISAs [6] |
| Microglia Differentiation Kits | Generating iPSC-derived microglia (iMGLs) | Protocol for iMGL generation and state manipulation [7] |
| 3D Multi-Electrode Arrays | Functional assessment of 3D neural networks | Custom 3D MEAs compatible with commercial systems (Multi Channel Systems MEA2100) [8] |
| Synaptosome Preparation Kits | Isolating synaptic fractions for pathway analysis | Synaptosome fractions for microglial state induction [7] |
| CRISPR/Cas9 Tools | Gene editing for isogenic controls or novel mutations | Creating isogenic APOE3/APOE4 lines; introducing fAD mutations [2] |
Applications in Drug Discovery and Therapeutic Development
iPSC-derived AD models are playing an increasingly important role in drug discovery, with several candidates identified through iPSC-based screening advancing to clinical trials [9]. These include bosutinib, ropinirole, and ezogabine for amyotrophic lateral sclerosis (ALS), demonstrating the broader applicability of iPSC-based screening for neurodegenerative diseases [9]. For AD specifically, iPSC models have been used to test γ-secretase modulators, Aβ aggregation inhibitors, and compounds targeting tau hyperphosphorylation [2].
The ability of iPSC models to recapitulate patient-specific pathophysiology enables a precision medicine approach to therapy development. For instance, a 2025 NIH report highlighted an epilepsy drug, levetiracetam, that may slow brain atrophy specifically in individuals without the APOE ε4 allele, demonstrating how treatments may work for some subpopulations but not others [1]. Similarly, the small molecule CT1812 shows promise for treating multiple types of dementia by displacing toxic protein aggregates at synapses [1].
High-throughput screening using iPSC-derived neurons and glia, sometimes combined with artificial intelligence approaches, represents the next frontier in AD drug discovery [9]. These platforms allow for the rapid testing of compound libraries on human cells with disease-relevant genetics, potentially accelerating the identification of novel therapeutic candidates.
iPSC-derived neuronal models have fundamentally transformed our approach to studying Alzheimer's disease pathogenesis. The recapitulation of core AD hallmarks—amyloid plaques and neurofibrillary tangles—in these human cell-based systems provides an unprecedented window into the molecular and cellular events driving neurodegeneration. While challenges remain in fully modeling the complexity of late-stage AD pathology in vitro, continued advancements in 3D culture systems, microglial incorporation, and functional assessment technologies are rapidly closing the gap between in vitro models and the human brain.
The application of these models to drug discovery holds particular promise for developing more effective, personalized therapies for this devastating disorder. As iPSC technology continues to evolve, it will undoubtedly play an increasingly central role in unraveling the mechanisms of Alzheimer's disease and developing interventions to combat it.
Alzheimer's disease (AD) represents a profound public health crisis, affecting millions worldwide without curative therapies. The disease is categorized into two primary forms: familial AD (FAD), a rare early-onset form caused by highly penetrant autosomal dominant mutations, and sporadic AD (sAD), a late-onset form with complex aetiologies involving polygenic risk and environmental factors [10]. Research into these distinct forms has been hampered by models that fail to recapitulate human-specific disease mechanisms, contributing to numerous failed clinical trials [11].
The advent of human induced pluripotent stem cell (iPSC) technology has revolutionized AD research by enabling the generation of patient-specific neural cells and tissues. This platform provides an unprecedented opportunity to model both FAD and sAD under physiologically relevant human genetic backgrounds [12]. For FAD, iPSCs allow precise dissection of pathogenic mechanisms stemming from known mutations, while for sAD, they facilitate exploration of the complex interplay between polygenic risk profiles and environmental influences [13]. This technical guide examines current methodologies, experimental paradigms, and applications of iPSC-based models for both FAD and sAD, providing a comprehensive resource for researchers and drug development professionals working at the intersection of genetics and neurodegenerative disease.
Genetic Architecture of FAD and sAD
The genetic underpinnings of FAD and sAD display distinct patterns of inheritance, risk loci, and molecular mechanisms, necessitating different modeling approaches.
Familial AD (FAD) Genetics
FAD follows an autosomal dominant inheritance pattern with nearly complete penetrance. It is primarily linked to mutations in three genes:
- APP (Amyloid Precursor Protein): Located on chromosome 21, with over 24 known pathogenic mutations that alter amyloid-beta production or aggregation properties [10].
- PSEN1 (Presenilin 1): Located on chromosome 14, with approximately 185 identified mutations that predominantly affect γ-secretase activity [10].
- PSEN2 (Presenilin 2): Located on chromosome 1, with at least 13 documented mutations that similarly dysregulate γ-secretase function [10].
A key unifying feature of FAD mutations is their common molecular phenotype: nearly all pathogenic variants in APP, PSEN1, and PSEN2 increase the Aβ42:Aβ40 ratio, promoting amyloid-beta aggregation and oligomerization [10].
Sporadic AD (sAD) Genetics
sAD exhibits a complex polygenic risk architecture influenced by numerous genetic variants with small effect sizes. The most significant genetic risk factor is the ε4 allele of APOE, which alone may account for up to 50% of sAD heritability [10]. Recent large-scale genome-wide association studies (GWAS) have identified over 70 genomic loci associated with sAD risk [14]. A 2025 multi-ancestry GWAS utilizing whole-genome sequencing data from diverse cohorts identified 16 novel AD loci, advocating for more genetically inclusive studies [14].
The emerging understanding of sAD genetics highlights the importance of polygenic risk scores (PRS) that aggregate the effects of many common variants. Research resources like the IPMAR Resource have established iPSC collections capturing extremes of global AD polygenic risk, including lines from high-risk late-onset AD, high-risk early-onset AD, and low-risk cognitively healthy controls [13].
Table 1: Key Genetic Features of FAD and sAD
| Feature | Familial AD (FAD) | Sporadic AD (sAD) |
|---|---|---|
| Heritability Pattern | Autosomal dominant | Polygenic, complex inheritance |
| Primary Genetic Drivers | Mutations in APP, PSEN1, PSEN2 | APOE ε4 allele + numerous risk loci |
| Age of Onset | Early-onset (<60 years) | Late-onset (≥65 years) |
| Proportion of Cases | ~5% | ~95% |
| Molecular Pathway | Altered Aβ42:Aβ40 ratio | Multiple pathways including lipid metabolism, immune response, endocytosis |
| iPSC Modeling Approach | Introduce known pathogenic mutations | Capture polygenic risk profiles; study gene-environment interactions |
iPSC Modeling Approaches for FAD and sAD
Establishing iPSC Models for FAD
FAD modeling leverages precise genome engineering to introduce specific pathogenic mutations into control iPSCs or utilizes iPSCs derived from FAD patients.
CRISPR/Cas9 Genome Editing: The CRISPR/Cas9 system enables targeted introduction of FAD-associated mutations into control iPSC lines. This approach involves:
- Design of sgRNAs targeting specific loci in APP, PSEN1, or PSEN2
- Delivery of CRISPR/Cas9 components alongside donor DNA templates containing desired mutations
- Selection and validation of successfully edited clones through sequencing and functional assays [15]
Isogenic Control Generation: A critical aspect of FAD modeling is creating isogenic control lines where the pathogenic mutation is corrected in patient-derived iPSCs, ensuring that observed phenotypes are mutation-specific rather than due to background genetic variation [15].
Advanced Modeling Strategies for sAD
sAD modeling requires more complex approaches to capture its polygenic nature:
Polygenic Risk Score Stratification: Donor selection based on calculated PRS allows establishment of iPSC cohorts representing extremes of genetic risk. The IPMAR Resource, for instance, includes 90 iPSC lines with extremes of global AD polygenic risk and 19 lines with complement pathway-specific genetic risk [13].
Multi-ancestry Cohort Representation: Recent studies emphasize the importance of including diverse genetic backgrounds in sAD modeling, as novel risk loci have been identified through inclusion of non-European ancestry individuals [14].
Environmental Challenge Models: sAD iPSC models can be exposed to various environmental risk factors, including Aβ42 oligomers to simulate amyloid stress or other triggers that might unmask latent genetic risks [16].
Experimental Platforms and Phenotypic Assays
2D Cortical Neuronal Cultures
Two-dimensional cortical neuronal cultures derived from FAD or sAD iPSCs provide a platform for high-content screening and mechanistic studies.
Functional Assays Using Microelectrode Arrays (MEA): Patterned iPSC-derived cortical neurons integrated with MEAs enable measurement of long-term potentiation (LTP), a cellular correlate of learning and memory. This system effectively models early-stage AD dysfunction without significant neuronal death [16].
LTP Impairment and Rescue: Studies demonstrate that Aβ42 oligomers significantly reduce LTP maintenance in human cortical neurons, and this impairment can be rescued by AD therapeutics including Donepezil, Memantine, Rolipram, and Saracatinib [16].
Protocol: MEA-based LTP Assay
- Differentiate iPSCs to cortical neurons using dual-SMAD inhibition
- Plate neurons on patterned MEAs to form defined circuits (≥8 weeks maturation)
- Record baseline neural activity and network properties
- Induce LTP using tetanic electrical stimulation (e.g., 100 Hz, 1s duration)
- Monitor post-tetanic potentiation for ≥1 hour
- Challenge system with Aβ42 oligomers (500 nM-1 μM) with/without therapeutic compounds
- Quantify LTP maintenance as percentage of baseline response [16]
3D Brain Organoid Models
Three-dimensional brain organoids provide more physiologically relevant environments for AD modeling by incorporating multiple neural cell types and complex cell-cell interactions.
Vascularized Neuroimmune Organoids: Advanced organoid systems now incorporate neurons, astrocytes, microglia, and blood vessels, creating comprehensive models that better mimic the brain's cellular ecosystem. These complex organoids can be exposed to AD patient brain extracts containing proteopathic seeds to induce multiple AD pathologies within four weeks [11].
Pathology Induction in sAD Organoids: Exposure of vascularized neuroimmune organoids to sAD brain extracts induces:
- Aβ plaque-like aggregates
- Tau tangle-like aggregates
- Neuroinflammation with elevated microglial synaptic pruning
- Synapse and neuronal loss
- Impaired neural network activity [11]
Protocol: Vascularized Neuroimmune Organoid Generation
- Differentiate iPSCs to neural progenitor cells (NPCs), primitive macrophage progenitors (PMPs), and vascular progenitors (VPs)
- Combine progenitors at optimized ratios (30,000 NPCs:12,000 PMPs:7,000 VPs)
- Allow self-assembly into 3D organoids in low-attachment plates
- Culture with mitogens (bFGF) for 5 days (proliferation stage)
- Transition to differentiation medium with IL-34 and VEGF for long-term maturation
- At day 30-40, challenge with sAD brain extracts (10% v/v) for 4 weeks
- Analyze pathological hallmarks via immunostaining and functional assays [11]
Table 2: Comparison of iPSC-based Experimental Platforms for AD Research
| Platform | Key Features | Applications | Limitations |
|---|---|---|---|
| 2D Cortical Cultures | Pure neuronal populations; amenable to high-content screening; MEA compatibility | Reductionist studies of neuronal vulnerability; electrophysiological characterization; HTS | Lacks cellular diversity; simplified connectivity |
| Conventional Brain Organoids | 3D architecture; multiple neural cell types; emergent network properties | Modeling neurodevelopment; compound screening; study of cell-cell interactions | Variable reproducibility; limited vascularization and immune components |
| Vascularized Neuroimmune Organoids | Includes neurons, astrocytes, microglia, and blood vessels; more complete brain mimicry | Modeling neuroinflammation; blood-brain barrier studies; sAD pathology induction | Technically challenging; longer maturation times; higher costs |
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Research Reagent Solutions for iPSC-based AD Modeling
| Reagent/Category | Specific Examples | Function/Application |
|---|---|---|
| CRISPR Components | Cas9 nuclease, sgRNAs targeting APP/PSEN1/PSEN2, HDR templates | Introduction or correction of FAD-associated mutations; creation of isogenic controls |
| Differentiation Reagents | Dual-SMAD inhibitors (LDN193189, SB431542), N2/B27 supplements, BDNF, GDNF | Directed differentiation of iPSCs to cortical neurons, astrocytes, or other neural lineages |
| 3D Culture Systems | Matrigel, laminin, recombinant IL-34, VEGF, bFGF | Support for organoid formation and maturation; promotion of vascularization and microglial development |
| Pathology Inducers | Synthetic Aβ42 oligomers, tau fibrils, AD patient brain extracts | Induction of AD-like pathologies in otherwise healthy models; challenge tests for resilience |
| Functional Assay Tools | Microelectrode arrays, calcium indicators, patch-clamp systems | Measurement of neuronal activity, synaptic function, and network properties |
| Therapeutic Compounds | Donepezil, Memantine, Rolipram, Saracatinib, Lecanemab | Validation of models through rescue experiments; drug screening applications |
Signaling Pathways in Alzheimer's Disease Pathogenesis
The following diagram illustrates key signaling pathways dysregulated in Alzheimer's disease, highlighting potential therapeutic targets:
Applications in Drug Discovery and Therapeutic Development
iPSC models of both FAD and sAD have become invaluable platforms for drug discovery and validation, addressing critical gaps in traditional preclinical models.
Target Identification and Validation
Genetic Risk Locus Functionalization: iPSC models enable functional validation of newly identified genetic risk factors. For instance, a 2024 study identified a novel genetic locus on chromosome 2p22.2 (between CYP1B1 and RMDN2) associated with tau deposition that accounts for 4.3% of tau PET variation - surpassing even APOE4's contribution (3.6%) [17]. iPSC models provide a platform to investigate the functional consequences of such loci.
Pathway Analysis: Proteomic studies of AD iPSC models have revealed disrupted AD-related pathways beyond the canonical amyloid and tau pathways, including complement system, oxidative stress response, and mitochondrial function [11].
Compound Screening and Validation
Phenotypic Screening Platforms: iPSC-derived models serve in high-content therapeutic screening for both target-based and phenotypic approaches. The MEA-based LTP platform has been validated with four AD therapeutics (Donepezil, Memantine, Rolipram, and Saracatinib), demonstrating rescue of Aβ42-induced LTP impairment [16].
Drug Efficacy Testing: Complex vascularized neuroimmune organoids have been used to test FDA-approved therapies like Lecanemab, showing significant reduction of amyloid burden following treatment [11].
Clinical Trial Predictive Value: Several clinical trials have been initiated based on iPSC research findings, including trials of bosutinib, ropinirole, and ezogabine for ALS, and WVE-004 and BII078 for ALS/FTD, demonstrating the translational potential of these models [9].
iPSC-based modeling of both FAD and sAD has fundamentally advanced our ability to study Alzheimer's disease in human-specific systems that recapitulate key genetic and pathological features. The distinct approaches required for each form - precise genetic engineering for FAD versus polygenic risk capture and environmental challenge for sAD - highlight the complementary value of these models.
Future developments will likely focus on increasing model complexity through improved organoid systems with better vascularization and inclusion of additional cell types, enhancing maturation to better model late-onset processes, and incorporating high-content multi-omics readouts for comprehensive phenotyping. The integration of artificial intelligence with iPSC-based screening is already emerging as a powerful approach for pattern recognition and predictive modeling [9].
As these technologies continue to evolve, iPSC models of FAD and sAD will play an increasingly central role in bridging the gap between genetic discoveries and functional pathophysiology, ultimately accelerating the development of effective therapeutics for this devastating disorder. The ongoing creation of large-scale, well-characterized iPSC resources capturing the spectrum of AD genetic risk promises to provide the research community with powerful tools to dissect disease mechanisms and identify novel therapeutic interventions.
Alzheimer's disease (AD) research is undergoing a paradigm shift, moving beyond the neuron-centric view to embrace the critical contributions of glial cells in disease pathogenesis. While historical research emphasized amyloid-β (Aβ) plaques and neurofibrillary tangles as the hallmark pathologies, recent genetic and molecular evidence reveals that non-neuronal cells play equally important roles in disease initiation and progression [18]. The emergence of human induced pluripotent stem cell (iPSC) technology has been instrumental in this reconceptualization, enabling researchers to model the complex cell-type-specific contributions to AD pathophysiology within a human context. iPSC-derived glial models provide unprecedented access to the human-specific aspects of neuroinflammation that cannot be adequately captured in rodent models [19].
The growing recognition that numerous AD risk genes identified through genome-wide association studies (GWAS) are predominantly expressed in glia has further highlighted these cells as essential therapeutic targets [18]. Technologies such as iPSC differentiation and CRISPR/Cas9 gene editing have converged to create powerful platforms for investigating glial functions in AD, allowing researchers to dissect the roles of microglia and astrocytes in neuroinflammatory processes with human-specific relevance [20]. This technical guide explores how these advanced iPSC-based models are reshaping our understanding of AD mechanisms and accelerating the development of novel therapeutic strategies.
The Neuroinflammatory Landscape in Alzheimer's Disease
Central Players: Microglia and Astrocytes
In the central nervous system (CNS), microglia and astrocytes serve as the primary mediators of neuroinflammation, engaging in complex, dynamic responses to disease-related insults [19]. Rather than existing in simple binary states (e.g., resting/activated), these cells exhibit a spectrum of functional phenotypes that vary based on specific contextual signals.
Microglia, the brain's resident immune cells, constitute 0.5-16.6% of the total brain cell population depending on brain region, sex, and developmental stage [19]. In AD, microglia respond to damage-associated molecular patterns (DAMPs) such as Aβ oligomers through pattern recognition receptors, triggering activation of NF-κB signaling pathways and subsequent release of pro-inflammatory cytokines including IL-1α, TNF, and C1q [19]. These microglial-derived factors stimulate astrocytes to acquire a reactive phenotype, establishing a feedforward loop of neuroinflammatory signaling.
Astrocytes, the most abundant CNS glial cell type (17-61% of total brain cells), perform diverse functions including neurotransmitter recycling, metabolic support, and blood-brain barrier maintenance [19]. In AD, astrocytes become reactive, characterized by increased expression of glial fibrillary acidic protein (GFAP) and morphological changes that correlate with patient clinical markers [21]. Reactive astrocytes contribute to AD pathogenesis through multiple mechanisms including disrupted glutamate homeostasis leading to excitotoxicity, impaired Aβ clearance, and secretion of inflammatory mediators that amplify neuroinflammatory responses [22].
Signaling Pathways in Glial-Driven Neuroinflammation
The following diagram illustrates key neuroinflammatory signaling pathways between microglia and astrocytes in Alzheimer's disease:
Figure 1: Neuroinflammatory Signaling in Alzheimer's Disease. This diagram illustrates how amyloid-β (Aβ) and damage-associated molecular patterns (DAMPs) activate microglial NF-κB signaling, triggering cytokine release that induces astrocyte reactivity and contributes to synaptic and neuronal damage.
iPSC-Derived Glial Models: Methodological Approaches and Technical Considerations
Differentiation Strategies for Microglia and Astrocytes
Current protocols for generating microglia and astrocytes from human iPSCs generally follow one of two approaches: directed differentiation recapitulating developmental processes, or direct conversion using transcription factor programming [23]. The directed differentiation method sequentially mimics in vivo developmental stages through carefully timed exposure to specific growth factors and small molecules, while direct conversion approaches induce glial fates through overexpression of key transcription factors.
Microglia Differentiation: Protocols typically involve first generating hematopoietic progenitor cells, then maturing these precursors into microglia-like cells using combinations of cytokines including IL-34, M-CSF, GM-CSF, and TGF-β to promote microglial identity [19]. The resulting iPSC-derived microglia (iMG) exhibit characteristic ramified morphology, express canonical microglial markers (P2RY12, TMEM119, IBA1), and display functional properties including phagocytosis, cytokine secretion, and chemotaxis [24].
Astrocyte Differentiation: Most protocols begin with neural induction of iPSCs to generate neural progenitor cells (NPCs), followed by astrocyte specification using CNTF, BMPs, or LIF, with maturation requiring extended culture periods (often 60-180 days) [23]. The resulting iPSC-derived astrocytes express standard markers (GFAP, S100β, EAAT1/GLAST) and demonstrate functional properties including glutamate uptake, calcium signaling, and inflammatory responsiveness [21].
Table 1: Key Differentiation Factors for iPSC-Derived Glial Cells
| Cell Type | Key Differentiation Factors | Maturation Time | Characteristic Markers |
|---|---|---|---|
| Microglia | IL-34, M-CSF, GM-CSF, TGF-β | 4-6 weeks | P2RY12, TMEM119, IBA1, CX3CR1 |
| Astrocytes | CNTF, BMP2/4, LIF, FGF2 | 8-26 weeks | GFAP, S100β, EAAT1/GLAST, EAAT2/GLT-1 |
Advanced Model Systems for Neuroinflammation Research
To better capture the complexity of CNS inflammation, researchers have developed increasingly sophisticated culture systems:
Monocultures: Single-cell type cultures enable reductionist studies of cell-autonomous responses but lack intercellular interactions [19].
Conditioned Media Transfer: This approach allows investigation of paracrine signaling without direct cell-cell contact by exposing one cell type to soluble factors secreted by another [19].
2D Co-culture Systems: Direct co-culture of multiple cell types (e.g., neurons with microglia or astrocytes) facilitates study of cell-contact-dependent signaling and bidirectional communication [19].
3D Organoid Systems: Cerebral organoids containing multiple CNS cell types self-organize into complex structures that more closely mimic the cellular diversity and organization of the developing brain [23].
Xenotransplantation Models: iPSC-derived human glia transplanted into mouse brains integrate into host circuits and adopt more mature phenotypes through exposure to the in vivo environment [19].
The following workflow diagram illustrates the process for establishing these diverse model systems:
Figure 2: Experimental Workflow for iPSC Glial Model Generation. This diagram outlines the differentiation of iPSCs into various neural lineages and the subsequent establishment of different model systems for neuroinflammation research.
Applications in Disease Modeling and Therapeutic Development
Modeling Patient-Specific Pathophysiology
iPSC-derived glial models demonstrate remarkable ability to recapitulate key features of individual patient pathophysiology. A compelling example comes from Rowland et al., who generated iPSC-derived astrocytes from AD patients stratified by levels of the inflammatory marker YKL-40 in cerebrospinal fluid [21]. Their research revealed that astrocyte morphological changes in response to Aβ oligomers correlated with patient clinical markers, with astrocytes from patients with lower CSF-YKL-40 levels and more protective APOE genotypes showing the greatest morphology changes [21]. This finding was subsequently verified using unbiased similarity learning approaches, highlighting how iPSC-derived glia capture biologically meaningful aspects of patient-specific disease phenotypes.
For modeling the complex genetics of sporadic AD, large-scale resources like the IPMAR Resource (iPSC Platform to Model Alzheimer's Disease Risk) have been developed, featuring over 100 iPSC lines capturing extremes of global AD polygenic risk as well as pathway-specific genetic risk (e.g., complement pathway) [13]. This collection includes lines from patients with high-risk early- and late-onset AD as well as low-risk cognitively healthy controls, many of whom have lived beyond the age of risk for disease development (>85 years) [13]. Such resources enable systematic investigation of how polygenic risk shapes glial contributions to AD pathogenesis.
High-Content Screening and Phenotypic Profiling
Advanced imaging and machine learning approaches are being leveraged to extract quantitative phenotypic information from iPSC-derived glial models. High-content imaging combined with deep learning algorithms enables unbiased quantification of morphological dynamics in glial cells following exposure to pathological insults [21]. These approaches can detect subtle, disease-relevant phenotypic changes that might be missed in conventional analyses, providing rich datasets for patient stratification and compound screening.
Table 2: Quantitative Assessments in iPSC-Glial Alzheimer's Models
| Parameter Measured | Experimental Approach | Key Findings |
|---|---|---|
| Morphological Changes | High-content imaging, deep learning | Astrocyte morphology changes reflect patient clinical markers and APOE genotype [21] |
| Cytokine Secretion | Multiplex immunoassays (Luminex), ELISA | Distinct inflammatory profiles in high-risk vs. low-risk genotypes |
| Phagocytic Activity | pH-sensitive fluorescent beads, Aβ uptake assays | Impaired Aβ clearance in APOE4 genotypes |
| Calcium Signaling | GCaMP imaging, Fluo-4 assays | Aberrant calcium dynamics in reactive astrocytes |
| Metabolic Function | Seahorse assays, mitochondrial staining | Bioenergetic deficits in disease-associated glia |
Therapeutic Screening and Cell Engineering
iPSC-derived glial models are accelerating therapeutic development through multiple approaches:
Drug Screening: Glial models enable screening of compounds targeting neuroinflammatory processes. For instance, researchers are testing whether epilepsy drugs like levetiracetam might be repurposed for AD based on findings of abnormal electrical activity in AD brains [1]. Preliminary results suggest this approach may slow brain atrophy in individuals without the APOE ε4 allele, highlighting the importance of genotype-stratified therapeutic responses [1].
Cell Therapy Engineering: CRISPR-engineered iPSC-derived microglia are being developed as innovative therapeutic delivery vehicles. A groundbreaking study demonstrated that iPSC-microglia engineered to express the Aβ-degrading enzyme neprilysin under control of a plaque-responsive promoter (CD9) could reduce multiple biochemical measures of pathology in AD mice [24]. Both regional and CNS-wide engraftment approaches reduced Aβ pathology, but only widespread microglial engraftment achieved significant reductions in plaque load, dystrophic neurites, and astrogliosis while preserving neuronal density in plaque-dense regions [24].
Gene Editing: CRISPR/Cas9 enables precise manipulation of AD-related genes in iPSC-derived glia to study disease mechanisms and potential therapeutic interventions. For example, editing of PSEN1 and PSEN2 in iPSC-derived cells has provided insights into how these mutations alter Aβ production and glial function [20]. Similar approaches targeting APOE have revealed allele-specific effects on Aβ clearance and neuroinflammatory responses [20].
The Scientist's Toolkit: Essential Research Reagents and Methodologies
Research Reagent Solutions
Table 3: Essential Research Reagents for iPSC-Glial Neuroinflammation Studies
| Reagent Category | Specific Examples | Research Application |
|---|---|---|
| Differentiation Factors | IL-34, M-CSF, GM-CSF, TGF-β, CNTF, BMP2/4 | Direct iPSC differentiation toward microglial or astrocytic lineages |
| Inflammatory Stimuli | Aβ oligomers, LPS, TNF-α, IL-1α/β, IFN-γ | Induction of neuroinflammatory responses in glial cultures |
| CRISPR Components | Cas9 nucleases, sgRNAs, HDR templates | Genetic engineering to introduce or correct AD-related mutations |
| Cell Surface Markers | Anti-CD11b, anti-TMEM119, anti-P2RY12 (microglia); anti-GFAP, anti-S100β (astrocytes) | Identification and purification of specific glial cell types |
| Cytokine Detection | Multiplex cytokine arrays, ELISA kits for TNF-α, IL-6, IL-1β | Quantification of inflammatory mediator secretion |
Protocol: Modeling Aβ-Induced Neuroinflammation in iPSC-Derived Glial Co-cultures
Experimental Workflow:
- Differentiate iPSCs to microglia and astrocytes using established protocols [23]
- Establish co-culture systems by seeding iMG and iAstrocytes in optimized ratios (typically 1:2 to 1:5 microglia:astrocytes)
- Treat with Aβ oligomers (500 nM-2 μM) for 24-72 hours to induce AD-relevant neuroinflammation
- Implement experimental readouts including:
- High-content imaging for morphological analysis
- Multiplex cytokine profiling of conditioned media
- RNA sequencing for transcriptomic analysis
- Immunocytochemistry for marker expression and localization
- Functional assays (phagocytosis, glutamate uptake)
Key Technical Considerations:
- Include appropriate controls (vehicle-treated, monotherapy conditions)
- Use isogenic iPSC lines when possible to control for genetic background effects
- Validate Aβ oligomer preparation size and aggregation state using western blot or TEM
- Consider temporal dynamics of inflammatory responses through time-course experiments
Future Directions and Concluding Perspectives
The integration of iPSC-derived glial models into Alzheimer's disease research has fundamentally expanded our understanding of disease mechanisms, highlighting neuroinflammation as a central driver of pathogenesis rather than merely a secondary consequence. The development of increasingly complex culture systems, including multi-cell type co-cultures and region-specific organoids, will further enhance the physiological relevance of these models [23]. Meanwhile, advanced gene editing technologies like CRISPR/Cas9 enable precise dissection of how specific genetic variants contribute to glial dysfunction in AD [20].
Looking forward, several key areas will shape the next generation of iPSC-glial research. Multi-omics integration (transcriptomics, proteomics, epigenomics) applied to iPSC-derived glia from genetically stratified donors will reveal novel molecular pathways governing neuroinflammatory responses [13]. High-throughput screening platforms using iPSC-derived glia will accelerate the identification of compounds that modulate neuroinflammation in patient-specific contexts [21]. Finally, engineered glial cell therapies represent a promising frontier, with recent studies demonstrating the potential of iPSC-derived microglia to deliver therapeutic proteins throughout the CNS in a pathology-responsive manner [24].
As these technologies mature, iPSC-derived glial models will increasingly guide therapeutic development for Alzheimer's disease, enabling precision medicine approaches that account for individual genetic backgrounds and disease subtypes. By faithfully capturing the critical contributions of microglia and astrocytes to neuroinflammation, these human cellular models provide an essential bridge between basic research and clinical translation in the ongoing effort to develop effective Alzheimer's therapeutics.
Within the broader thesis on the mechanisms of Alzheimer's disease (AD) using induced pluripotent stem cell (iPSC) models, this whitepaper provides an in-depth technical examination of three critical downstream pathological pathways: synaptic dysfunction, oxidative stress, and axonal transport deficits. The advent of iPSC technology has revolutionized AD research by enabling the generation of patient-specific neural cells that recapitulate human disease pathophysiology in vitro [23] [25] [26]. These human cellular models are particularly valuable for investigating downstream events in AD pathogenesis, as they overcome limitations of animal models that often fail to fully mimic human disease processes [27]. iPSC-derived neurons from both familial and sporadic AD patients have been demonstrated to faithfully model key disease phenotypes, including elevated amyloid-β (Aβ) levels, TAU hyperphosphorylation, increased oxidative stress sensitivity, and impaired neuronal function [6] [26]. This technical guide summarizes current methodologies, quantitative findings, and experimental approaches for investigating these downstream pathways using iPSC-based AD models, providing researchers with practical tools for advancing therapeutic discovery.
Pathological Hallmarks in iPSC-Derived AD Models
iPSC-derived neural models successfully recapitulate the major pathological hallmarks of Alzheimer's disease, providing a human-relevant platform for investigating disease mechanisms and therapeutic interventions. The table below summarizes key pathological phenotypes observed in iPSC-based AD models.
Table 1: Key Pathological Phenotypes in iPSC-Derived Alzheimer's Disease Models
| Pathological Hallmark | iPSC Model Findings | Significance | References |
|---|---|---|---|
| Amyloid-β Pathology | Increased Aβ1–40 and Aβ1–42 levels; Elevated Aβ1–42/Aβ1–40 ratio in fAD | Recapitulates amyloid cascade hypothesis; Differentiates fAD from sAD | [6] [26] |
| TAU Hyperphosphorylation | Increased phosphorylation at multiple sites (Thr231, Ser396, etc.); GSK3B activation | Models neurofibrillary tangle formation; Links amyloid and tau pathology | [6] |
| Oxidative Stress Sensitivity | Enhanced vulnerability to amyloid oligomers and peroxide | Demonstrates increased oxidative stress susceptibility in AD neurons | [6] |
| Axonal Spheroid Formation | Plaque-associated axonal spheroids (PAAS) with disrupted conduction | Correlates with AD severity; disrupts neural circuits | [28] |
| Synaptic Dysfunction | Altered presynaptic (SNAP25, GAP43) and postsynaptic (NRGN) markers | Strongest structural correlate of cognitive deficits | [27] |
iPSC models have been particularly valuable for studying both familial (fAD) and sporadic (sAD) forms of Alzheimer's disease. Neurons derived from fAD patients (typically carrying PSEN1 mutations) and sAD patients show remarkably similar phenotypes, including increased phosphorylation of TAU protein at all investigated phosphorylation sites and higher levels of extracellular Aβ1–40 and Aβ1–42 [6]. However, the significantly increased Aβ1–42/Aβ1–40 ratio—a pathological marker of fAD—is observed primarily in fAD samples, highlighting both shared and distinct pathological mechanisms between AD forms [6]. These findings demonstrate that iPSC technology is suitable for modeling both fAD and sAD, providing a platform for developing novel treatment strategies for these conditions.
Synaptic Dysfunction in AD iPSC Models
Mechanisms and Pathological Features
Synaptic dysfunction represents the strongest structural correlate of cognitive deficits in Alzheimer's disease and is considered a central feature of AD pathogenesis [27]. iPSC-based studies have revealed that synaptic damage in AD involves multiple mechanisms, including local accumulation of Aβ around plaques causing localized damage to synapses of nearby neurons, thereby disturbing local neuronal networks [27]. Research using iPSC-derived neurons has demonstrated that high Aβ concentrations in and around plaques trigger damage to glutamatergic transmission and nearby synapses by targeting postsynaptic glutamatergic receptors in a calcium-dependent manner [27].
The analysis of cerebrospinal fluid biomarkers in AD patients has revealed specific alterations in synaptic proteins that can be modeled in iPSC systems. These include changes in presynaptic markers such as SNAP25 and GAP43 (involved in synaptic plasticity) and the postsynaptic marker neurogranin (NRGN, involved in long-term potentiation) [27]. These synaptic alterations are believed to contribute to memory dysfunction and disrupted default-mode network activity in the AD brain.
Experimental Assessment Protocols
Table 2: Methodologies for Assessing Synaptic Dysfunction in iPSC-Derived Neurons
| Method | Key Parameters | Technical Considerations | Applications | |
|---|---|---|---|---|
| Immuno-cytochemistry | Presynaptic (SNAP25, GAP43) and postsynaptic (PSD95, NRGN) marker localization and quantification | Requires high-resolution imaging (confocal/STED); Quantitative analysis of puncta density and size | Mapping spatial distribution of synaptic proteins; Assessing synapse density | [27] |
| Electro-physiology | Spontaneous postsynaptic currents; Network activity using MEA; Long-term potentiation (LTP) | Requires mature neuronal cultures (>60 days); Technical expertise in patch clamping | Functional assessment of synaptic transmission and plasticity | [29] |
| CSF Biomarker Analysis | Levels of SNAP25, GAP43, NRGN in conditioned media | Sensitivity of detection methods; Correlation with clinical biomarkers | Translational validation; Drug response assessment | [27] |
| Live-cell Imaging | Synaptic vesicle recycling (FM dyes); Calcium imaging | Temporal resolution; Phototoxicity considerations | Real-time monitoring of synaptic function and dynamics | [28] |
Oxidative Stress Mechanisms
Oxidative Stress in AD Pathogenesis
Oxidative stress represents a significant downstream pathway in Alzheimer's disease pathology, with iPSC-derived AD neurons demonstrating enhanced sensitivity to oxidative insults. Studies have shown that both fAD- and sAD-derived neurons exhibit elevated sensitivity to oxidative stress induced by amyloid oligomers or peroxide [6]. This increased vulnerability reflects the compromised antioxidant defense mechanisms and heightened oxidative damage observed in the AD brain.
The relationship between oxidative stress and other AD pathological features is complex and bidirectional. Aβ accumulation can induce oxidative stress by promoting mitochondrial dysfunction and generating reactive oxygen species (ROS). Conversely, oxidative stress can accelerate amyloidogenesis by enhancing β- and γ-secretase activities while inhibiting α-secretase processing of APP [6]. Additionally, oxidative stress can promote tau hyperphosphorylation by activating various kinases, including GSK3β, further connecting these pathological pathways.
Assessment Methodologies
The protocol for evaluating oxidative stress vulnerability in iPSC-derived AD neurons typically involves exposing mature neuronal cultures (60+ days of differentiation) to specific oxidative stressors, such as hydrogen peroxide or amyloid oligomers, followed by assessment of cell viability and metabolic function [6]. Additional measures include:
- ROS Detection: Using fluorescent probes like CM-H2DCFDA to measure intracellular ROS levels
- Mitochondrial Function Assessment: Utilizing JC-1 staining or TMRM to measure mitochondrial membrane potential
- Antioxidant Defense Evaluation: Quantifying expression and activity of antioxidant enzymes (SOD, catalase, glutathione peroxidase)
- Lipid Peroxidation Measurement: Assessing levels of malondialdehyde (MDA) or 4-hydroxynonenal (4-HNE)
These approaches allow researchers to quantify the increased oxidative stress sensitivity observed in AD-derived neurons and evaluate potential therapeutic compounds that might mitigate this vulnerability.
Axonal Transport Deficits
Plaque-Associated Axonal Spheroids (PAAS)
A recent breakthrough in understanding axonal pathology in AD comes from the identification and characterization of plaque-associated axonal spheroids (PAAS) using iPSC models and proteomic approaches [28]. These dystrophic neurites are found around amyloid deposits in Alzheimer's disease, where they impair axonal electrical conduction, disrupt neural circuits, and correlate with AD severity [28]. Hundreds of axons near individual plaques develop enlarged spheroid-like structures that contain enlarged, enzyme-deficient endolysosomal vesicles and autophagosomes [28].
A proximity labeling proteomics approach has been developed to characterize the molecular composition of PAAS in postmortem AD human brains and iPSC-derived models [28]. This method utilizes PLD3, an endolysosomal protein highly abundant within PAAS, as a protein bait for proximity labeling. The approach involves sequential incubation with primary antibody against PLD3 and an HRP-conjugated secondary antibody, followed by a peroxidation reaction with H2O2 and Biotin-XX-Tyramide, resulting in robust biotinylation of proteins within PAAS [28]. Proteomic analysis of these structures has revealed abnormalities in key biological processes, including protein turnover, cytoskeleton dynamics, and lipid transport.
mTOR Pathway in Axonal Pathology
Notably, the PI3K/AKT/mTOR pathway—which regulates these processes—was found to be activated in spheroids, with phosphorylated mTOR levels in spheroids correlating with AD severity in humans [28]. This discovery has significant therapeutic implications, as mTOR inhibition in iPSC-derived neurons and mice ameliorated spheroid pathology [28]. The following diagram illustrates the key signaling pathways involved in axonal spheroid formation and potential therapeutic targets:
Diagram Title: mTOR Pathway in Axonal Spheroid Formation
Experimental Protocols for iPSC-Based AD Research
Neural Differentiation Workflow
The investigation of downstream pathways in AD using iPSC models requires robust differentiation protocols to generate relevant neural cell types. The following diagram outlines a standardized workflow for generating AD-relevant neural cells from iPSCs:
Diagram Title: iPSC Neural Differentiation Workflow
The standard neural induction protocol typically employs dual SMAD inhibition using Noggin and SB431542 to efficiently direct differentiation toward neural fates [23]. This approach generates PAX6+ neural cells competent of rosette formation within approximately 11 days [23]. Neural rosettes can then be isolated and expanded as neural stem cells (NSCs), which can subsequently be differentiated into mature neuronal and glial cell types.
High-Content Phenotypic Screening
For drug discovery applications, high-content phenotypic screening platforms have been developed using iPSC-derived neurons. These typically involve:
- Longitudinal Live-cell Imaging: Automated imaging systems to monitor neuronal health and degeneration over time [29]
- Metabolic Assays: Measurement of cellular viability and metabolic function (e.g., MTT, ATP assays)
- Immunocytochemical Analysis: High-throughput staining and imaging of key pathological markers (Aβ, p-TAU, synaptic proteins)
- Multi-electrode Array (MEA): Functional assessment of neuronal network activity [29]
- Transcriptomic Analysis: RNA sequencing to evaluate pathway alterations and drug responses
These approaches have been validated in large-scale studies, such as a recent screen of 100 sporadic ALS iPSC lines (relevant to neurodegenerative disease mechanisms generally), which demonstrated the ability to recapitulate key disease phenotypes and identify potential therapeutic compounds [29].
Research Reagent Solutions
The table below provides a comprehensive list of essential research reagents and their applications in iPSC-based AD research, compiled from the cited literature:
Table 3: Essential Research Reagents for iPSC-Based Alzheimer's Disease Research
| Reagent/Category | Specific Examples | Function/Application | References |
|---|---|---|---|
| Neural Induction Agents | Noggin, SB431542, LDN-193189 | Dual SMAD inhibition for efficient neural differentiation | [23] |
| Neural Maturation Factors | BDNF, GDNF, NT-3, cAMP, Ascorbic acid | Promote neuronal maturation and synaptic development | [23] [29] |
| Cell Type Markers | PAX6 (NSCs), Tuj1 (neurons), MAP2 (mature neurons), GFAP (astrocytes) | Identification and validation of specific neural cell types | [23] [29] |
| Pathological Antibodies | Anti-Aβ, Anti-phospho-TAU (Thr231, Ser396, etc.), Anti-GSK3B | Detection of AD pathological hallmarks | [28] [6] |
| Pathway Modulators | mTOR inhibitors (e.g., Rapamycin), GSK3B inhibitors, BACE1 inhibitors | Mechanistic studies and therapeutic target validation | [28] [6] |
| Proteomic Tools | PLD3 antibody, Biotin-XX-Tyramide, Streptavidin beads | Proximity labeling for subcellular proteomics | [28] |
| Vital Dyes & Reporters | CM-H2DCFDA (ROS), JC-1 (mitochondrial potential), Calcium indicators | Functional assessment of oxidative stress and signaling | [28] [6] |
iPSC-based models have emerged as powerful tools for investigating downstream pathways in Alzheimer's disease, including synaptic dysfunction, oxidative stress, and axonal transport deficits. These patient-derived cellular models recapitulate key aspects of AD pathology and provide a human-relevant platform for mechanistic studies and drug discovery. The integration of advanced techniques such as proximity labeling proteomics, high-content screening, and 3D organoid cultures has enabled unprecedented insights into the molecular mechanisms driving AD pathogenesis. Furthermore, the identification of potentially reversible pathways, such as mTOR activation in axonal spheroids, provides new hope for therapeutic interventions. As iPSC technology continues to evolve, it will undoubtedly play an increasingly central role in unraveling the complex mechanisms of Alzheimer's disease and developing effective treatments for this devastating disorder.
Uncovering Novel Therapeutic Targets through Genomic and Transcriptomic Analysis of iPSC-Derived Brain Cells
Alzheimer's disease (AD) remains a devastating neurodegenerative disorder with no cure, characterized by progressive cognitive decline and pathological hallmarks including amyloid-beta (Aβ) plaques and neurofibrillary tangles [20] [18]. The failure of countless therapeutics developed in traditional rodent models has highlighted the critical need for human-relevant model systems that better recapitulate human disease pathophysiology [18]. The advent of human induced pluripotent stem cell (iPSC) technology has revolutionized AD research by enabling the generation of patient-specific brain cells, including neurons, astrocytes, microglia, and more complex 3D organoid systems [12] [11] [18].
Integrating genomic and transcriptomic analyses with these iPSC-derived models provides unprecedented opportunities to uncover novel therapeutic targets by directly linking human genetic risk factors to functional cellular and molecular phenotypes [30] [31]. This approach allows researchers to move beyond the limitations of familial AD models and investigate the complex polygenic nature of sporadic AD, which accounts for over 95% of cases [30] [11]. The ability to capture individual genetic backgrounds while manipulating specific risk genes through CRISPR/Cas9 genome editing further enables precise dissection of disease mechanisms [20] [15]. This technical guide outlines how these powerful technologies are being integrated to advance our understanding of AD pathogenesis and identify novel therapeutic intervention points.
Polygenic Risk Score Stratification in iPSC Cohorts
The complex genetic architecture of sporadic Alzheimer's disease necessitates iPSC resources that capture the polygenic nature of disease risk. The IPMAR Resource (iPSC Platform to Model Alzheimer's Disease Risk) represents a significant advancement, comprising 109 iPSC lines selected from extremes of global AD polygenic risk and complement pathway-specific genetic risk [30].
Table 1: IPMAR Resource Composition for Genomic Risk Modeling
| Risk Category | Donor Diagnosis | Number of Lines | Mean Polygenic Risk Score | Key Characteristics |
|---|---|---|---|---|
| Global AD Polygenic Risk | Late-onset AD (LOAD) | 34 | 2.2 ± 0.5 SD | Age of onset: 72 ± 6 years; 56% female |
| Global AD Polygenic Risk | Early-onset AD (EOAD) | 29 | 2.1 ± 0.4 SD | Age of onset: 51 ± 3 years; 55% female |
| Global AD Polygenic Risk | Cognitively healthy controls | 27 | -1.9 ± 0.4 SD | 59% female; elderly controls (>85 years) |
| Complement Pathway Risk | LOAD | 9 | 2.4 ± 0.3 SD | Age of onset: 71 ± 6 years; 78% female |
| Complement Pathway Risk | Cognitively healthy controls | 10 | -1.9 ± 0.2 SD | 70% female |
This resource enables researchers to investigate how aggregated genetic risk across the genome, quantified as polygenic risk scores (PRS), influences cellular phenotypes in human-derived neurons and glia [30]. The PRS approach aggregates information from all identified genetic variants known to impact overall AD risk, providing a personalized estimate of an individual's genetic susceptibility with 84% accuracy in predicting disease development [30].
APOE Isoform-Specific Modeling
Beyond polygenic risk scores, specific high-risk genetic factors can be modeled using isogenic iPSC lines. The APOE ε4 allele represents the strongest genetic risk factor for late-onset AD, with APOE2 considered protective and APOE3 neutral [31] [18]. Researchers have generated iPSC-derived microglia with different APOE genotypes (APOE2, APOE3, APOE4, and APOE-knockout) transplanted into mouse brains to study microglial-autonomous effects [31].
Table 2: Key Genomic Targets in Alzheimer's Disease iPSC Modeling
| Gene/Pathway | Risk Allele/Variant | Associated AD Risk | Primary Cellular Functions | iPSC Modeling Approaches |
|---|---|---|---|---|
| APOE | APOE4 (vs. APOE3) | 3-fold (heterozygous) 15-fold (homozygous) | Lipid transport, Aβ clearance, immune regulation | Isogenic lines, xenotransplantation, transcriptomics |
| APOE | APOE2 (vs. APOE3) | Protective | Lipid transport, Aβ clearance, immune regulation | Isogenic lines, xenotransplantation, transcriptomics |
| APP | Multiple FAD mutations | Highly penetrant early-onset AD | Aβ production, synaptic function | CRISPR correction, pathogenic mutation introduction |
| PSEN1/PSEN2 | Multiple FAD mutations | Highly penetrant early-onset AD | γ-secretase activity, Aβ generation | CRISPR knockout, pathogenic mutation introduction |
| Complement Pathway | Multiple SNPs from GWAS | Modest individual effects, significant in aggregate | Immune clearance, synaptic pruning | Polygenic risk score stratification, pathway analysis |
RNA-seq and ATAC-seq analyses of these microglia revealed widespread transcriptomic and epigenomic differences dependent on APOE genotype, with the most significant differences observed between the protective APOE2 and risk-associated APOE4 isoforms [31]. APOE4 microglia showed impaired proliferation, migration, and immune responses, while APOE2 microglia exhibited enhanced phagocytic capabilities and unique vitamin D receptor signaling [31].
Transcriptomic Profiling of iPSC-Derived Brain Cells
Single-Cell RNA Sequencing of Complex Co-culture Systems
Advanced 3D model systems now enable transcriptomic profiling of multiple cell types in physiologically relevant contexts. A vascularized neuroimmune organoid model containing neurons, astrocytes, microglia, and blood vessels has been developed to study AD pathogenesis in a more complex human system [11]. When exposed to brain extracts from sporadic AD patients, these organoids develop multiple AD pathologies within four weeks, including Aβ plaque-like aggregates, tau tangle-like aggregates, neuroinflammation, synaptic loss, and impaired neural network activity [11].
Single-nuclei RNA sequencing (snRNA-seq) of similar 3D neurosphere models has revealed how cellular interactions shape transcriptional responses to chronic amyloidosis [32]. In neurospheres containing neurons, astrocytes, and microglia, chronic exposure to Aβ oligomers induced significant transcriptional changes that were profoundly influenced by the presence of microglia [32]. Specifically, microglia were essential for the upregulation of AD-associated genes in astrocytes, including APOE, CLU, LRP1, and VIM, highlighting the critical role of intercellular signaling in shaping disease-relevant transcriptomic responses [32].
Figure 1: Experimental workflow for transcriptomic analysis of iPSC-derived 3D neurospheres under chronic amyloidosis conditions.
Subcellular Proteomics of Disease-Associated Structures
Beyond transcriptomics, subcellular proteomic approaches have been developed to characterize the molecular composition of specific pathological structures in AD. Proximity labeling techniques using enzymes like horseradish peroxidase (HRP) conjugated to antibodies against proteins enriched in pathological compartments enable proteomic profiling of these structures [28]. For example, targeting phospholipase D3 (PLD3), highly enriched in plaque-associated axonal spheroids (PAAS), has revealed the proteome of these dystrophic neurites [28].
This approach identified 821 proteins representing the PAAS proteome, with enrichment in biological processes including protein turnover, cytoskeleton dynamics, and lipid transport [28]. Notably, the PI3K/AKT/mTOR pathway was activated in spheroids, and phosphorylated mTOR levels correlated with AD severity in humans [28]. Furthermore, mTOR inhibition in iPSC-derived neurons and mouse models ameliorated spheroid pathology, validating this pathway as a therapeutic target identified through subcellular proteomics [28].
Advanced 3D Model Systems for Pathological Recapitulation
Vascularized Neuroimmune Organoids
Recent advancements in 3D culture systems have enabled the development of more physiologically relevant models that incorporate multiple cell types affected in AD. Vascularized neuroimmune organoids contain human neurons, microglia, astrocytes, and blood vessels, providing a comprehensive system to study cellular interactions in AD pathogenesis [11]. These organoids exhibit functional blood vessels with endothelial cells and pericytes, as demonstrated by staining for CD31 (endothelial marker) and PDGFRβ (pericyte marker) [11].
When exposed to postmortem brain extracts from sporadic AD patients, these organoids develop multiple AD pathologies within four weeks, including:
- Aβ plaque-like aggregates and tau tangle-like aggregates
- Neuroinflammation with activated microglia
- Elevated microglial synaptic pruning
- Synapse and neuronal loss
- Impaired neural network activity [11]
This model also demonstrates utility for therapeutic testing, as treatment with Lecanemab, an FDA-approved anti-Aβ antibody, significantly reduced amyloid burden in AD brain extract-exposed organoids [11].
Chronic Amyloidosis Neurosphere Models
Another 3D approach uses iPSC-derived neurospheres containing neurons and astrocytes, with optional addition of microglia, to study cellular responses to chronic amyloidosis [32]. In this system, microglia efficiently phagocytose Aβ and significantly reduce neurotoxicity, mitigating amyloidosis-induced oxidative stress and neurodegeneration [32]. The neuroprotective effects conferred by microglia were associated with unique gene expression profiles in astrocytes and neurons, including several known AD-associated genes such as APOE [32].
Table 3: Essential Research Reagents for iPSC-Based Alzheimer's Disease Modeling
| Reagent/Cell Type | Specification | Function in Model System | Key Characterization Markers |
|---|---|---|---|
| Neural Progenitor Cells (NPCs) | iPSC-derived, PAX6+/NESTIN+ | Foundation for neuronal and astrocyte differentiation | PAX6, NESTIN, SOX1, SOX2 |
| Primitive Macrophage Progenitors (PMPs) | iPSC-derived, CD235+/CD43+ | Source for microglia in co-culture systems | CD235, CD43, PU.1 |
| Vascular Progenitors (VPs) | iPSC-derived, CD31+ | Forms vascular structures in 3D organoids | CD31, PDGFRβ, Collagen IV |
| Brain Organoid Media | Supplemented with IL-34, VEGF, neurotrophic factors | Supports maturation of neuronal, microglial, and vascular components | N/A (culture medium) |
| Aβ Oligomers | Synthetic Aβ1-42 | Induces chronic amyloidosis pathology in 3D models | N/A (experimental treatment) |
This model provides a platform to study the protective functions of microglia and their role in shaping astrocyte and neuronal responses to Aβ, highlighting how different cell types interact to modify disease progression [32].
Therapeutic Target Validation and Screening
Target Identification Through Multi-Omic Integration
The integration of genomic and transcriptomic data from iPSC-derived brain cells has identified numerous potential therapeutic targets for AD. Key pathways emerging from these analyses include:
- mTOR Signaling: Subcellular proteomics of axonal spheroids identified mTOR activation as a key process in PAAS formation, with mTOR inhibition reducing spheroid pathology in iPSC-derived neurons and mouse models [28].
- APOE-Related Pathways: Transcriptomic profiling of APOE isoform-specific microglia revealed impaired proliferation, migration, and immune responses in APOE4 microglia, while APOE2 microglia showed enhanced phagocytic capability and unique vitamin D receptor signaling [31].
- Complement System: iPSC lines stratified by complement pathway-specific polygenic risk enable investigation of how genetic variation in this pathway influences neuroimmune function and AD susceptibility [30].
- Inflammatory Response: snRNA-seq of 3D neurosphere models revealed microglia-dependent upregulation of inflammatory genes in astrocytes, including APOE, CLU, LRP1, and VIM, in response to Aβ exposure [32].
Figure 2: Integrated workflow for therapeutic target discovery using iPSC models and multi-omic profiling.
Experimental Validation Approaches
Once potential therapeutic targets are identified through genomic and transcriptomic analyses, several experimental approaches enable validation:
CRISPR/Cas9 Genome Editing: CRISPR/Cas9 enables precise genetic modifications in key AD-related genes, such as APP, PSEN1, PSEN2, and APOE, in iPSC lines [20] [15]. This approach allows researchers to correct pathogenic mutations or introduce risk variants in isogenic backgrounds to study their specific effects on molecular and cellular phenotypes [20].
Pharmacological Screening: iPSC-derived neurons, glia, and 3D organoids provide human-relevant platforms for drug screening. For example, mTOR inhibition was shown to ameliorate axonal spheroid pathology in both iPSC-derived neurons and mouse models [28]. Similarly, Lecanemab treatment reduced amyloid burden in vascularized neuroimmune organoids exposed to AD brain extracts [11].
Functional Assays: Comprehensive functional characterization including electrophysiology, calcium imaging, phagocytosis assays, and neuronal activity measurements enables correlation of transcriptomic findings with functional outcomes [28] [32]. For instance, in 3D neurosphere models, microglia were shown to protect against Aβ-induced dysfunction of neuronal activity and oxidative stress [32].
Several clinical trials have been initiated based on findings from iPSC research, including trials of bosutinib, ropinirole, and ezogabine for amyotrophic lateral sclerosis (ALS), demonstrating the translational potential of this approach [33]. Similar strategies are being applied to AD, with the goal of accelerating therapeutic development.
The integration of genomic and transcriptomic analyses with iPSC-derived brain models has transformed our ability to identify and validate novel therapeutic targets for Alzheimer's disease. These human-relevant systems capture the genetic complexity of AD and enable detailed investigation of disease mechanisms in precisely defined cellular contexts. As these technologies continue to advance—with improvements in 3D model complexity, single-cell multi-omic profiling, and genome editing—they hold considerable promise for delivering the next generation of AD therapeutics that have proven so elusive in traditional model systems. The ongoing development of large, genetically stratified iPSC resources, combined with advanced analytical approaches, will be critical to realizing the potential of personalized medicine for this devastating disorder.
Building Better Brain Models: iPSC Differentiation, High-Content Screening, and Drug Discovery Platforms
Protocols for Differentiating iPSCs into Functional Cortical Neurons and Motor Neurons
The development of robust protocols for differentiating induced pluripotent stem cells (iPSCs) into specific neuronal subtypes represents a cornerstone of modern neurological disease modeling, particularly for Alzheimer's disease (AD). Patient-derived iPSCs retain the individual's complete genetic background, enabling researchers to create human neuronal models that recapitulate disease-specific pathologies in vitro [34]. For AD research, the ability to generate cortical neurons—which are vulnerable to amyloid-beta and tau pathology—and motor neurons—relevant for understanding circuit dysfunction and axonal transport deficits—provides powerful platforms for investigating disease mechanisms and screening therapeutic candidates [12] [28]. This technical guide details established methodologies for generating these functionally mature neuronal populations, with specific consideration for their application in AD research.
Cortical Neuron Differentiation Protocols
Cortical neurons derived from human iPSCs provide essential models for studying AD-related pathologies, including amyloid plaque formation, tau hyperphosphorylation, and synaptic dysfunction observed in the cerebral cortex.
Defined Cortical Neuron Differentiation with Dual SMAD and Wnt Inhibition
A highly efficient 14-day protocol generates cortical neural stem cells (NSCs) using a defined, small molecule-based approach [35]. This method employs dual-SMAD inhibition combined with Wnt/β-Catenin inhibition to direct cells toward a dorsal cortical fate.
Key Protocol Steps:
- Neural Induction (Days 1-10): Culture iPSCs in Neural Induction Medium containing SB431542 (TGF-β inhibitor), LDN193189 (BMP inhibitor), and XAV939 (Wnt inhibitor).
- Neural Maintenance (Days 10-14): Passage cells and culture in Neural Maintenance Medium to promote NSC expansion and neural rosette formation.
- Terminal Differentiation: Plate NSCs on poly-L-ornithine/laminin-coated surfaces in neuronal differentiation media containing BDNF, GDNF, cAMP, and ascorbic acid for 4-6 weeks to obtain mature cortical neurons [35].
This protocol consistently yields NSCs expressing PAX6, NESTIN, and FOXG1, with subsequent differentiation producing cortical neurons positive for CTIP2, a layer V cortical marker [36] [35].
Phenotypic Maturation Model for Functional Cortical Neurons
Hickman and colleagues developed a phenotypic model that yields functionally mature cortical networks capable of long-term potentiation (LTP) without astrocyte co-culture [36]. The critical advancement lies in the extended maturation period and careful characterization.
Maturation Timeline and Characteristics:
- Day 21: Cultures contain immature neurons co-expressing excitatory (VGlut1) and inhibitory (GABA) markers.
- Day 40: Cultures exhibit purely excitatory phenotypes with sole VGlut1 expression and no detectable glial cells [36].
- Functional Validation: Day 40 neurons demonstrate sustained LTP for over 1 hour following high-frequency stimulation, whereas Day 21 neurons require GABAA receptor blockade with picrotoxin to achieve LTP [36].
Table 1: Key Markers During Cortical Neuron Maturation
| Time Point | Neuronal Markers | Glial Markers | Functional Properties |
|---|---|---|---|
| Day 21 | β-III-tubulin+, VGlut1+, GABA+ | GFAP- | LTP only with picrotoxin |
| Day 40 | β-III-tubulin+, VGlut1+, CTIP2+ | GFAP- | Sustained LTP for 1 hour |
| Day 56+ | Synaptophysin+, NMDAR1+, GluR1+ | GFAP- | Spontaneous synaptic activity |
Simplified Protocol for Electrophysiologically Mature Networks
A simplified differentiation approach generates electrophysiologically mature cortical neuronal networks without specialized media or astrocyte co-culture [37]. This method produces cultures with a consistent 60:40 ratio of neurons and astrocytes arising from common forebrain neural progenitors.
Functional Characteristics:
- Resting Membrane Potential: -58.2 ± 1.0 mV
- Action Potential Amplitude: 66.5 ± 1.3 mV
- Synaptic Activity: 74% of neurons exhibit spontaneous synaptic activity (amplitude: 16.03 ± 0.82 pA; frequency: 1.09 ± 0.17 Hz) [37]
This system demonstrates that functional maturity can be achieved through common progenitor differentiation rather than requiring exogenous astrocyte co-culture.
Motor Neuron Differentiation Protocols
Motor neuron differentiation protocols typically employ ventral patterning strategies to specify spinal cord identity, creating valuable models for investigating axonal pathology and transport deficits relevant to AD and other neurodegenerative diseases.
Small Molecule-Based Motor Neuron Differentiation
Motor neuron differentiation recapitulates developmental signaling pathways using small molecules to direct rostrocaudal and dorsoventral patterning [38].
Key Developmental Signals Recapitulated:
- Dorsoventral Patterning: Sonic hedgehog (SHH) activation via purmorphamine (PMA) or SAG specifies ventral progenitor identity.
- Rostrocaudal Patterning: Retinoic acid (RA) and Wnt signaling promote caudal spinal cord identity.
- Neural Induction: Dual-SMAD inhibition (SB431542 + LDN193189) directs neural commitment [38].
Standard 32-Day Protocol: The Answer ALS project optimized a 32-day direct motor neuron differentiation protocol based on dual-SMAD inhibition and ventral patterning [39]. This protocol has been successfully applied to differentiate over 400 iPSC lines from controls and ALS patients, demonstrating its robustness across large sample sets.
Characterization Results:
- Cultures contain heterogeneous cell populations including SMI-32+/TUBB3+ neurons and S100B+ astrocyte-like cells.
- ISL1+ motor neurons represent approximately 14-17% of total cells, with variations observed between sex and disease status [39].
- Male ALS samples showed significantly higher percentages of ISL1+ motor neurons compared to controls [39].
Table 2: Motor Neuron Differentiation Efficiency and Markers
| Marker | Expression in Differentiated Cultures | Cell Type Identified |
|---|---|---|
| ISL1 | 14.1% ± 0.8% (control), 17.0% ± 0.4% (ALS) | Spinal motor neurons |
| NKX6.1 | Enriched in male ALS samples | Motor neuron progenitors |
| TUBB3 | >60% of cells | General neuronal population |
| S100B | Present from day 18, increases by day 32 | Astrocyte-like cells |
Advanced Motor Neuron Modeling with Microfluidics and Co-culture
Recent advancements incorporate microfluidic devices to model axon-specific pathology and co-culture with skeletal muscle to create functional neuromuscular junctions [38]. These approaches are particularly relevant for investigating AD-related axonal spheroids, which disrupt electrical conduction and contribute to circuit dysfunction [28].
The Scientist's Toolkit: Essential Research Reagents
Table 3: Essential Reagents for iPSC Neuronal Differentiation
| Reagent | Function | Example Applications |
|---|---|---|
| SB431542 | TGF-β/Activin A/Nodal inhibitor; dual-SMAD component | Neural induction in cortical and motor neuron protocols [35] |
| LDN193189 | BMP receptor inhibitor; dual-SMAD component | Neural induction in cortical and motor neuron protocols [36] [35] |
| XAV939 | Wnt/β-catenin inhibitor; promotes cortical fate | Dorsal forebrain patterning in cortical differentiation [35] |
| Purmorphamine (PMA) | Sonic hedgehog pathway agonist; ventral patterning | Motor neuron specification [38] |
| Retinoic Acid (RA) | Rostrocaudal patterning; caudalization | Spinal motor neuron differentiation [38] |
| BDNF/GDNF | Trophic factors supporting neuronal survival | Neuronal maturation in both cortical and motor neuron protocols [36] [37] |
| cAMP/Ascorbic Acid | Promotes neuronal maturation and survival | Terminal differentiation in cortical neurons [36] [37] |
Signaling Pathways and Workflows
The following diagrams illustrate the key signaling pathways and experimental workflows for directing iPSC differentiation toward cortical and motor neuron fates.
Cortical Neuron Differentiation Signaling Pathway
Cortical Neuron Differentiation Pathway
Motor Neuron Differentiation Signaling Pathway
Motor Neuron Differentiation Pathway
Experimental Workflow for Alzheimer's Disease Modeling
iPSC Modeling Workflow for Alzheimer's Disease
Application to Alzheimer's Disease Research
iPSC-derived cortical and motor neurons provide critical human systems for investigating AD mechanisms and treatments. Recent research has leveraged these models to study axonal spheroids (dystrophic neurites) found around amyloid plaques in AD patients [28]. These spheroids disrupt axonal conduction, impair neuronal networks, and correlate with AD severity.
A multidisciplinary approach combining subcellular proteomics of human postmortem AD brains with iPSC-derived cortical neurons identified activation of the PI3K/AKT/mTOR pathway within axonal spheroids [28]. Notably, mTOR inhibition in iPSC-derived neurons ameliorated spheroid pathology, highlighting the utility of these models for identifying and validating therapeutic targets.
For drug discovery, iPSC-derived neuronal models from AD patients and controls have been used in high-throughput screening platforms. Several clinical trials based on iPSC research have emerged, including those for bosutinib, ropinirole, and ezogabine [33]. The integration of artificial intelligence with iPSC-based screening further enhances the drug discovery pipeline [33].
The continued refinement of protocols for differentiating iPSCs into cortical and motor neurons has established robust, reproducible platforms for modeling Alzheimer's disease mechanisms. The defined methodologies presented here—incorporating specific small molecule inhibitors, growth factors, and maturation timelines—enable researchers to generate functionally mature human neuronal networks that recapitulate key aspects of AD pathology. These systems now serve as invaluable tools for deciphering disease mechanisms, identifying novel therapeutic targets, and screening potential treatments, ultimately advancing our understanding and treatment of this devastating neurodegenerative disorder.
The study of Alzheimer’s disease (AD) has been hampered by the profound limitations of traditional model systems. Animal models, particularly mouse models that overexpress familial AD (FAD) mutant proteins, have been the cornerstone of preclinical research for decades yet have consistently failed to predict therapeutic efficacy in human patients [18]. This translational gap highlights a critical disconnect between rodent biology and human pathophysiology, particularly for a disease characterized by complex cellular interactions and lengthy pathological progression. The advent of induced pluripotent stem cell (iPSC) technology has revolutionized our approach by enabling the generation of patient-specific neural cells that carry the individual's complete genetic background [40]. However, conventional two-dimensional (2D) monocultures fail to recapitulate the intricate three-dimensional architecture and multicellular interactions of the human brain.
The integration of 3D brain organoids with sophisticated tri-culture systems represents a paradigm shift in AD modeling. Brain organoids are three-dimensional multicellular structures derived from pluripotent stem cells (PSCs) that self-organize to simulate in vivo brain regions to some extent [41]. When combined with tri-culture approaches that incorporate neurons, astrocytes, and microglia – the core cellular components of the "tripartite synapse" – these systems provide unprecedented opportunities to investigate the cell-type-specific contributions to AD pathogenesis in a human-derived context [42] [18]. This technical guide explores the development, application, and analysis of these advanced model systems within the framework of iPSC-based Alzheimer's disease research.
Core Principles: Brain Organoids and Tri-culture Systems
3D Brain Organoids: From Simple Spheroids to Complex Assembloids
Brain organoids represent a significant advancement over traditional 2D cultures by reproducing aspects of endogenous tissue architecture, cellular diversity, and network functionality [41]. Two primary methodologies have emerged for their generation: self-patterning approaches that leverage the innate self-organization potential of PSCs, and patterned protocols that use extrinsic morphogens to direct development toward specific brain regions [43]. The self-patterning method, pioneered by Lancaster and Knoblich, involves embedding pluripotent stem cell aggregates in extracellular matrix (ECM) to generate so-called "cerebral organoids" containing multiple brain regions [41] [43]. In contrast, patterned approaches apply precise sequences of signaling molecules to generate region-specific organoids resembling cortical, hippocampal, hypothalamic, or midbrain tissues [41].
Table 1: Brain Organoid Types and Their Applications in Alzheimer's Disease Research
| Organoid Type | Generation Method | Key Features | AD Modeling Applications |
|---|---|---|---|
| Cerebral Organoids | Self-patterning via ECM embedding [43] | Multiple brain regions; recapitulate early neurodevelopment | Studying early pathological events; network-level dysfunction |
| Region-Specific Organoids | Patterned using morphogen cocktails [41] | Specific brain regions (cortex, hippocampus); enhanced reproducibility | Region-specific vulnerability; pathway-specific pathology |
| Assembloids | Fusion of region-specific organoids [43] [42] | Model circuit formation between brain regions | Circuit-based spread of pathology; tau propagation studies |
| Vascularized Organoids | Co-culture with endothelial cells or incorporation of vascular progenitors [41] | Presence of vessel-like structures; improved nutrient exchange | Blood-brain barrier dysfunction; neurovascular unit in AD |
The limitations of simple organoid systems have prompted the development of increasingly complex models. Organoid fusion approaches create "assembloids" that model interactions between different brain regions, such as cortical-striatal or cortical-thalamic circuits, enabling the study of network dysfunction and pathological spread in AD [43]. Similarly, vascularization strategies enhance organoid maturation and viability while providing models for investigating blood-brain barrier dysfunction, a critical aspect of AD pathophysiology [41].
Tri-culture Systems: Recapitulating the Neuro-Glial-Vascular Unit
While neurons are undoubtedly central to AD pathogenesis, genetic studies have increasingly highlighted the crucial roles of glial cells in disease initiation and progression [18]. The tri-culture system, incorporating neurons, astrocytes, and microglia, reconstitutes the "tripartite synapse" and enables the investigation of neuroinflammation and glial contributions to neurodegeneration [42]. This system more realistically mimics the neuroinflammatory response in vivo, allowing a better understanding of how cellular crosstalk influences AD pathology [42].
Astrocytes contribute to AD pathogenesis through multiple mechanisms, including altered neurotransmitter recycling, impaired metabolic support, and dysfunctional phagocytosis of amyloid-β [18]. Microglia, the resident immune cells of the brain, are involved in synaptic pruning and clearance of protein aggregates, with numerous AD risk genes highly expressed in these cells [18]. The inclusion of both cell types in tri-culture systems enables researchers to model their synergistic effects on neuronal function and survival in AD.
Integrated Methodologies: Protocol for Generating AD-specific Tri-culture Organoids
Generation of iPSC-Derived Brain Organoids
The following protocol outlines the generation of cortical organoids, which are particularly relevant for AD research due to the vulnerability of cortical regions to AD pathology.
Day 0-6: Embryoid Body Formation and Neural Induction
- Culture human iPSCs (healthy controls or AD patients) in essential 8 medium until 70-80% confluent.
- Dissociate cells with Accutase and seed 9,000 cells per well in 96-well U-bottom low-attachment plates in neural induction medium supplemented with 50µM ROCK inhibitor [43].
- Centers should form embryoid bodies (EBs) within 24 hours.
- On day 2, replace medium with fresh neural induction medium without ROCK inhibitor.
- On day 6, EBs should have increased in size and show smooth, defined edges.
Day 7-30: Patterned Cortical Differentiation and Expansion
- Transfer EBs to 24-well low-attachment plates in cortical differentiation medium containing TGF-β/NODAL inhibitor (SB431542, 10µM) and BMP inhibitor (LDN193189, 100nM) to promote forebrain fate [41] [43].
- On day 10, embed EBs in Matrigel droplets (15-20 µL per EB) and transfer to 6-well plates with cortical differentiation medium.
- From day 15 onward, maintain organoids in cerebral organoid medium on an orbital shaker (60-80 rpm) to enhance nutrient exchange [43].
- Change medium every 3-4 days.
Day 30-90: Maturation and Long-term Culture
- Continue culture in cerebral organoid medium with reduced growth factors.
- For enhanced maturation, transfer day 45 organoids to sliced culture format or use spinning bioreactors to improve oxygen and nutrient penetration [41].
- Organoids develop cortical layer organization, functional synapses, and electrophysiological activity by 3 months [41].
Table 2: Key Media Formulations for AD-Relevant Brain Organoids
| Medium Type | Base Medium | Key Additives | Function in Organoid Development |
|---|---|---|---|
| Neural Induction | DMEM/F12 + N2 Supplement | NEAA, Heparin, Dual-SMAD inhibitors | Directs pluripotent cells toward neural lineage |
| Cortical Differentiation | DMEM/F12 + N2 & B27 Supplements | FGF2 (20ng/mL), EGF (20ng/mL) | Promotes forebrain and cortical identity |
| Cerebral Organoid | DMEM/F12 + B27 Supplement (without Vitamin A) | Insulin (2.5 µg/mL), BDNF (20ng/mL) | Supports general neuronal maturation and diversity |
| Maturation | Neurobasal + B27 Supplement | BDNF (20ng/mL), GDNF (10ng/mL), cAMP (250µM) | Enhances neuronal complexity and synaptic function |
Establishing the Tri-culture System
The integration of glial cells can be achieved through two primary approaches: direct differentiation within the organoid or incorporation of pre-differentiated glial cells.
Method A: Endogenous Glial Differentiation
- Maintain cortical organoids for extended periods (>3 months) to allow for the spontaneous differentiation of astrocytes and oligodendrocytes from neural progenitors [41].
- Microglia can be generated by adding microglia-promoting factors (IL-34, M-CSF, TGF-β) to the medium from day 30 onward [43].
- This method maintains the endogenous developmental trajectory but offers less control over glial cell ratios.
Method B: Incorporation of Pre-differentiated Glial Cells
- Differentiate astrocytes and microglia separately from the same iPSC line using established protocols.
- For astrocytes: Differentiate iPSCs to neural progenitor cells, then to glial progenitors using FGF2 and EGF, followed by astrocyte maturation with CNTF and BMP4 [18].
- For microglia: Differentiate iPSCs through hematopoietic progenitors using VEGF, SCF, and IL-3, then mature with IL-34, M-CSF, and TGF-β [18].
- At day 45 of organoid culture, dissociate the glial cells and add them to the organoid culture at a ratio of approximately 1:10 (microglia:neurons) and 1:3 (astrocytes:neurons) [42].
- Allow 7-14 days for glial integration and network formation.
Technical Considerations and Optimization Strategies
Enhancing Organoid Maturation and Viability
Long-term culture is essential for modeling late-onset diseases like AD but presents challenges including interior hypoxia and cell death. Several strategies have been developed to address these limitations:
- Sliced Organoid Culture: After 45 days, neocortical organoids can be sliced (150-200µm thickness) using a vibratome, which reduces inner hypoxia, diminishes cell death, and sustains neurogenesis, mimicking the embryonic human neocortex at the third trimester of gestation [41].
- Bioreactor Systems: Spinning bioreactors or orbital shakers improve nutrient and oxygen exchange, enabling organoids to reach more advanced developmental stages [43].
- Vascularization: Co-culture with endothelial cells or incorporation of mesodermal progenitors can promote the formation of vessel-like structures that enhance nutrient delivery and extend organoid lifespan [41] [42].
Reproducibility and Quality Control
The inherent heterogeneity of organoid systems presents challenges for reproducible research. The following approaches can enhance experimental consistency:
- Standardized Differentiation Protocols: Use precisely defined media and timing for each differentiation stage.
- Morphological Clustering: Image-based analysis of colony morphology can quantify and monitor culture status, enabling the identification of optimal differentiation conditions [44].
- Molecular Characterization: Regular assessment of regional markers (e.g., FOXG1 for forebrain, OTX2 for midbrain, PAX6 for cortical progenitors) ensures appropriate patterning.
- Functional Validation: Electrophysiological measurements (multi-electrode arrays, patch clamping) confirm neuronal maturity and network activity.
Applications in Alzheimer's Disease Research and Drug Discovery
Modeling AD Pathophysiology
Integrated brain organoid-tri-culture systems enable the investigation of multiple aspects of AD pathology:
- Aβ Pathology: Organoids generated from FAD-iPSCs or CRISPR-edited to express APP or PSEN1 mutations recapitulate key features of AD, including increased Aβ42/Aβ40 ratios and amyloid plaque-like aggregates after extended culture [18]. The tri-culture system allows assessment of cell-type-specific responses to Aβ and its clearance by microglia and astrocytes.
- Tau Pathology: Although tau aggregation develops more slowly, organoids containing neurons with MAPT mutations or the APOE4 genotype can model tau hyperphosphorylation and tangle-like formations [18]. The tri-culture system enables investigation of tau spreading between cells.
- Neuroinflammation: The inclusion of microglia and astrocytes facilitates the study of neuroinflammatory processes, including cytokine secretion, phagocytic activity, and their impact on neuronal health [42] [18].
- Genetic Risk Factors: Isogenic iPSC lines with modifications in AD risk genes (e.g., TREM2, CD33, APOE) can be used to investigate their cell-type-specific functions in a more physiological context [18].
Drug Discovery and Therapeutic Screening
iPSC-based models are increasingly being utilized in drug discovery pipelines for neurodegenerative diseases, with several candidates identified through iPSC screening advancing to clinical trials [9]. The enhanced physiological relevance of tri-culture organoid systems makes them particularly valuable for:
- Target Validation: Testing hypotheses about cell-type-specific drug targets in a human-relevant system.
- Compound Screening: Medium-to-high throughput screening of compound libraries for effects on AD-relevant phenotypes.
- Toxicity Assessment: Evaluating cell-type-specific toxicities that might not be apparent in simpler models.
- Personalized Medicine: Using patient-derived organoids to identify individualized therapeutic responses.
Advanced Imaging and Analysis Techniques
The complexity of 3D organoid systems demands sophisticated imaging and analysis approaches. High-content imaging (HCI) combined with artificial intelligence (AI)-based analysis pipelines enables quantitative assessment of complex phenotypes [45].
Figure 1: High-content imaging and analysis workflow for 3D organoids
High-Content Imaging Modalities
- Confocal Microscopy: Essential for optical sectioning of 3D samples, with water immersion objectives providing improved resolution for thick tissues [45] [46].
- Multi-photon Microscopy: Enables deeper imaging penetration for larger organoids.
- Live-cell Imaging: Time-lapse imaging tracks dynamic processes like neurite outgrowth, cell migration, and calcium signaling [47].
- Super-resolution Microscopy: Techniques such as STED or SIM resolve subcellular structures below the diffraction limit, enabling visualization of synaptic details [45].
AI-Based Image Analysis
Machine learning approaches are revolutionizing organoid image analysis:
- Nuclei Segmentation: U-Net architectures can accurately segment individual nuclei in phase contrast images, enabling label-free tracking of cell division and migration [47].
- Phenotypic Classification: Convolutional neural networks classify organoid regions, cell types, and pathological features [45].
- Morphological Analysis: AI algorithms quantify complex morphological parameters (volume, surface area, shape metrics) across thousands of organoids [46].
- Multi-parametric Analysis: Integrated analysis of multiple features (cell number, marker intensity, spatial distribution) enables comprehensive phenotypic profiling [45].
The Scientist's Toolkit: Essential Research Reagents and Platforms
Table 3: Essential Research Reagents and Platforms for Organoid Research
| Category | Specific Examples | Function/Application | Considerations for AD Research |
|---|---|---|---|
| Stem Cell Sources | Patient-derived iPSCs, Isogenic CRISPR-edited lines | Provide genetically defined starting material | APOE genotype; PSEN1/2/APP mutations; TREM2 variants |
| Extracellular Matrix | Matrigel, Synthetic hydrogels (e.g., alginate, PEG) | Provide 3D structural support for organoids | Composition affects Aβ aggregation and diffusion |
| Patterning Molecules | SMAD inhibitors (SB431542, LDN193189), Wnt agonists/antagonists, SHH agonists (SAG, purmorphamine) | Direct regional specification of organoids | Region-specific vulnerability (entorhinal cortex, hippocampus) |
| Maturation Factors | BDNF, GDNF, NT-3, cAMP | Enhance neuronal maturity and synaptic function | Critical for late-onset AD phenotype manifestation |
| Cell Type Markers | MAP2 (neurons), GFAP (astrocytes), IBA1 (microglia), OLIG2 (oligodendrocytes) | Characterize cellular composition | Assess cell-type-specific pathology in tri-cultures |
| Pathology Markers | Aβ antibodies (6E10, 4G8), p-tau (AT8, PHF1), Synaptophysin | Quantify AD-relevant pathology | Enable tracking of disease progression in vitro |
| Imaging Platforms | Confocal high-content systems (ImageXpress Confocal HT.ai), Spinning disk confocals | 3D imaging of organoids | Must accommodate long-term live imaging for longitudinal studies |
| Analysis Software | ImageJ, CellProfiler, IMARIS, IN Carta | Quantify morphology, intensity, and spatial relationships | Machine learning capabilities enhance pathological feature detection |
The integration of 3D brain organoids with tri-culture systems represents a transformative approach for modeling the complex multicellular pathophysiology of Alzheimer's disease. These advanced platforms bridge critical gaps between traditional animal models and human biology, offering unprecedented opportunities to study cell-type-specific contributions to disease mechanisms, screen therapeutic candidates in a human-relevant system, and advance personalized medicine approaches for this devastating disorder.
As the field progresses, key challenges remain, including further enhancing organoid maturation to better recapitulate adult brain physiology, improving reproducibility through standardized protocols, and integrating additional cell types such as vascular components to model the complete neuro-glial-vascular unit. The continued refinement of these integrated systems, coupled with advanced imaging and computational analysis approaches, promises to accelerate our understanding of Alzheimer's disease and the development of effective therapeutic strategies.
Functional phenotyping of neuronal networks is essential for understanding the mechanisms of neurological diseases, including Alzheimer's disease (AD). Among various technologies, multi-electrode array (MEA) has emerged as a powerful platform for non-invasive, long-term monitoring of neuronal activity in vitro. This technical guide details how MEA-based assays can be applied to study key functional phenotypes—particularly neuronal hyperexcitability, long-term potentiation (LTP), and network synchronization—within the context of human induced pluripotent stem cell (iPSC) models of AD. Impairment of LTP, the biological substrate for learning and memory, is a prominent feature in many neurological disorders, and establishing robust methods to assess it in human iPSC-derived neurons is crucial for disease modeling and drug development [48].
The advent of iPSC technology has provided unique opportunities to study AD using patient-specific neurons that retain the individual's complete genetic background. This is particularly valuable for modeling common, polygenic forms of AD, which account for the vast majority of cases. Large-scale iPSC resources, such as the IPMAR resource which includes lines from over 100 donors selected for extremes of AD polygenic risk, now enable researchers to investigate how genetic predisposition influences neuronal network function and synaptic plasticity [30]. By combining these physiologically relevant models with MEA technology, researchers can bridge the gap between molecular pathology and functional network deficits observed in AD.
MEA Technology and Network Activity Phenotypes
Multi-electrode array technology enables simultaneous recording of extracellular action potentials from dozens to hundreds of sites across a neuronal network. When applied to iPSC-derived neuronal cultures, this allows for the quantification of several key functional metrics over weeks or months, capturing the development and maturation of network activity.
Key Functional Metrics Quantifiable by MEA:
- Firing Rate: The average rate of action potentials per electrode, reflecting overall network excitability.
- Bursting Activity: Short periods of high-frequency firing followed by periods of silence, indicative of local network synchronization.
- Network Bursts (NBs): Synchronous bursts of action potentials across multiple electrodes, representing large-scale network synchronization.
In healthy iPSC-derived neuronal networks, such as commercially available iPSC-derived dopaminergic (DA) neurons in co-culture with astrocytes (e.g., iDopa/iAstro cultures), consistent baseline network activity typically emerges within 3–4 weeks after plating. Pharmacological characterization of these networks confirms their functional integrity: NB frequency is eliminated by the AMPA/kainate receptor blocker CNQX, strongly reduced by the NMDAR blocker AP5, and increased by the GABAA receptor blocker picrotoxin, demonstrating the balanced excitation and inhibition within the network [48].
In disease models, such as those for Amyotrophic Lateral Sclerosis (ALS), patient-derived motor neuron networks show clear signatures of network dysfunction, including increased firing rates and bursting activity [49]. This neuronal hyperexcitability is considered an early event in the neurodegenerative cascade. Similarly, aberrant network activity is a hypothesized functional phenotype in AD patient iPSC-derived neurons, making MEA a critical tool for detecting functional deficits and evaluating potential therapeutic interventions.
Measuring Long-Term Potentiation (LTP) with MEA
Background and Significance
Long-term potentiation is a persistent, activity-dependent strengthening of synaptic efficacy that is widely considered the cellular correlate of learning and memory. LTP impairment is a common feature in pre-clinical models of many neurological disorders, including Alzheimer's disease, Parkinson's disease, and autism [48]. Traditional methods to induce LTP, such as high-frequency electrical stimulation, activate only a small number of synapses. Chemical LTP (cLTP) protocols, however, can induce potentiation across entire neuronal networks simultaneously, making them particularly suitable for MEA platforms and enabling the detection of associated molecular changes [48].
Chemical LTP (cLTP) Induction Protocols
Two primary cLTP induction protocols have been successfully applied to iPSC-derived neuronal cultures on MEAs, each acting through distinct but convergent molecular pathways.
Table 1: Chemical LTP (cLTP) Induction Protocols for iPSC-Derived Neurons
| Induction Method | Key Reagents | Proposed Mechanism of Action | Duration of Potentiation | Applicable Cell Types |
|---|---|---|---|---|
| Forskolin/Rolipram (FSK/ROL) | Forskolin (adenylate cyclase activator), Rolipram (PDE inhibitor) | Increases cAMP levels, activates PKA, and triggers downstream LTP machinery [48] | Up to 72 hours after drug washout [48] | iPSC-derived midbrain dopaminergic and cortical neurons [48] |
| Tetraethylammonium (TEA) | Tetraethylammonium (K+ channel blocker) | Blocks K+ channels, leading to membrane depolarization, Ca2+ influx, and activation of CaMKII [49] | Several hours to days (observed in ALS motor neurons) [49] | iPSC-derived motor neurons (e.g., ALS models) [49] |
The FSK/ROL-mediated cLTP in iPSC-derived dopaminergic neurons induces a long-lasting increase in firing rate and NB frequency. This potentiation is notably largely independent of the NMDAR and is instead partially dependent on Brain-Derived Neurotrophic Factor (BDNF) and its receptor, TrkB [48]. Furthermore, this protocol triggers rapid phosphorylation of CREB and induction of canonical activity-regulated gene expression, confirming the activation of molecular pathways essential for late-phase LTP [48].
In ALS patient-derived motor neurons, TEA-induced cLTP has been shown to have therapeutic potential, tempering motor neuron hyperactivity and stabilizing spine density. This was accompanied by a temporary reduction in firing rate and bursting, and proteomic analysis revealed a restoration of dysregulated synaptic and metabolic pathways [49].
Experimental Workflow for cLTP-MEA Assay
A standardized workflow is essential for obtaining reliable and reproducible cLTP data. The following steps outline a typical cLTP-MEA assay.
Quantitative Data Presentation and Analysis
Robust quantification of MEA data before and after cLTP induction is critical for evaluating synaptic plasticity. The following parameters should be calculated and compared between baseline and post-induction periods.
Table 2: Key Quantitative Metrics for cLTP Analysis on MEA
| Parameter | Description | Interpretation in cLTP Context | Example Change Post-cLTP |
|---|---|---|---|
| Mean Firing Rate (Hz) | Average number of spikes per second per electrode. | Reflects overall increase in network excitability. | Increase of 50-150% [48] |
| Network Burst (NB) Frequency | Rate of occurrence of synchronous network bursts. | Indicates enhanced large-scale synchronization. | Increase of 50-200% [48] |
| NB Duration | Average length of network bursts. | May reflect stability of sustained synchronous activity. | Variable |
| Number of Spikes per NB | Mean spike count within each network burst. | Suggests strengthening of synaptic connections within the network. | Increase |
| Inter-Burst Interval | Time interval between consecutive network bursts. | Shorter intervals indicate increased network drive. | Decrease |
Data from the FSK/ROL cLTP assay on iDopa cultures showed that the potentiation could last for up to 72 hours after drug washout, allowing for the study of the transcription-dependent late phase of LTP (L-LTP) [48]. This long-lasting effect is crucial for investigating the molecular mechanisms that stabilize synaptic strengthening.
The Scientist's Toolkit: Research Reagent Solutions
Table 3: Essential Reagents and Tools for MEA-based Functional Phenotyping
| Item | Function/Description | Example Use Case |
|---|---|---|
| iPSC-Derived Neurons | Patient-specific or commercially available differentiated neurons. | iDopa neurons (midbrain dopaminergic) for Parkinson's research; cortical or motor neurons for AD/ALS studies [48] [49]. |
| 48-well MEA Plates | Multi-electrode array plates facilitating higher-throughput pharmacological interrogation. | Axion BioSystems plates used for cLTP assays on iPSC-derived dopaminergic networks [48]. |
| cLTP Inducers: FSK/ROL | Chemical cocktail to induce cAMP-dependent LTP across the network. | Forskolin (e.g., 50 µM) and Rolipram (e.g., 0.1 µM) applied for 15-30 minutes [48]. |
| cLTP Inducer: TEA | Potassium channel blocker to induce depolarization-dependent LTP. | Tetraethylammonium (e.g., 25 mM) applied for a defined period [49]. |
| Receptor Antagonists | Pharmacological blockers to dissect mechanisms (e.g., AP5, CNQX, PTX). | Used to characterize the contributions of NMDAR, AMPAR, and GABAAR to network activity [48]. |
| BDNF/TrkB Pathway Modulators | Recombinant BDNF or TrkB inhibitors to probe pathway-specific contributions. | Investigating partial BDNF-dependence of cLTP in dopaminergic neurons [48]. |
| Analysis Software | Software for spike sorting, burst detection, and network analysis. | Axion Biosystems' Axis Navigator and NeuroExplorer, or custom scripts in Python/MATLAB. |
Integrating MEA-based functional phenotyping with human iPSC-derived models represents a powerful approach for investigating the complex mechanisms of Alzheimer's disease and other neurodegenerative disorders. The ability to quantitatively measure neuronal hyperexcitability, network synchronization, and most importantly, synaptic plasticity via LTP, provides a direct readout of circuit function in a patient-specific context. The protocols detailed herein, particularly the cLTP assays, enable researchers to not only characterize disease phenotypes but also to probe the underlying molecular pathways and test potential therapeutic strategies aimed at restoring synaptic function and network balance. As iPSC resources that capture the polygenic nature of AD become more widespread [30], the application of these functional assays will be critical for translating genetic findings into mechanistic understanding and effective treatments.
High-Throughput and High-Content Drug Screening in iPSC-Derived Neuronal Libraries
Human induced pluripotent stem cell (iPSC)-derived neuronal models are revolutionizing Alzheimer's disease (AD) research by providing genetically accurate, patient-specific systems for drug discovery. AD drug development has been plagued by high clinical trial failure rates, partly due to reliance on animal models that fail to fully recapitulate human disease pathophysiology. iPSC technology enables the generation of patient-derived neurons and glia that retain the unique genetic makeup of the individual, including the complex polygenic risk architecture underlying sporadic AD, which constitutes over 95% of cases [50] [30]. These models allow researchers to study disease mechanisms in human cells relevant to AD, such as cortical neurons, and to screen for therapeutics in a human genetic context early in the drug discovery pipeline.
The integration of high-throughput screening (HTS) and high-content imaging (HCI) technologies with iPSC-derived neuronal libraries has created powerful platforms for identifying and validating potential AD therapeutics. These approaches enable the assessment of compound effects across thousands of conditions while capturing multiparametric data on complex neuronal phenotypes at single-cell resolution. This technical guide examines current methodologies, experimental protocols, and analytical frameworks for implementing high-throughput and high-content drug screening in iPSC-derived neuronal libraries specifically for Alzheimer's disease research.
Generation and Characterization of iPSC-Derived Neuronal Libraries
Library Generation Strategies
Establishing a well-characterized iPSC library is the foundational step for meaningful drug screening. For Alzheimer's disease research, libraries should encompass the genetic diversity of the patient population, including both familial and sporadic AD cases. Two recent large-scale initiatives demonstrate different approaches to library generation:
The IPMAR Resource (iPSC Platform to Model Alzheimer's Disease Risk) comprises 109 iPSC lines selected to capture extremes of polygenic risk for common AD. This includes 63 lines from patients with high global polygenic risk scores (34 late-onset AD, 29 early-onset AD), 27 from cognitively healthy controls with low polygenic risk, and 19 lines selected specifically for complement pathway-specific genetic risk (9 AD patients, 10 controls) [30]. This strategy enables researchers to stratify patients based on genetic risk profiles and identify subtype-specific therapeutic responses.
The SALS Motor Neuron Library represents another approach, featuring iPSCs from 100 sporadic ALS patients, 11 suspected monogenic cases, and 25 healthy controls. All lines underwent rigorous quality control including genomic integrity, pluripotency confirmation, and trilineage differentiation potential testing [29]. This library captures clinical heterogeneity with donors classified into ALS subtypes (lower motor neuron-predominant, classic ALS, upper motor neuron-predominant) and extensive clinical data including site of onset, age of onset, disease progression rates, and survival times [29].
Table 1: iPSC Library Configurations for Neurodegenerative Disease Research
| Library Characteristic | IPMAR AD Resource | SALS Motor Neuron Library |
|---|---|---|
| Total Lines | 109 | 136 |
| AD/ALS Cases | 72 | 111 |
| Healthy Controls | 37 | 25 |
| Selection Basis | Polygenic risk scores | Clinical heterogeneity |
| Genetic Characterization | PRS, APOE ε alleles, complement pathway SNPs | Whole-genome sequencing, pathogenic variants |
| Associated Data | Clinical, longitudinal, genetic | Clinical progression, survival, onset data |
Neuronal Differentiation Protocols
Robust, standardized differentiation protocols are essential for generating reproducible neuronal cultures for screening. Most protocols follow a sequential approach manipulating developmental signaling pathways:
Neural Progenitor Cell-Based Differentiation utilizes dual-SMAD inhibition (BMP and TGFβ signaling inhibition) to selectively block endoderm and mesodermal fates, enriching for neural ectoderm lineage progenitors [50]. iPSCs can form embryoid body-like aggregates followed by neural rosette structures that are precursors to NPCs. Cortical precursors can be further enriched using fluorescence-activated cell sorting for specific surface markers (CD184-positive, CD133-positive, CD271-negative) [50]. These NPCs can be stably propagated and differentiated into astrocytes, oligodendrocytes, and functional neurons.
Direct Small Molecule-Based Differentiation approaches, such as the protocol adapted for the SALS motor neuron screening, use a five-stage process with extensively optimized maturation conditions capable of discriminating between healthy and diseased motor neurons [29]. The protocol generates high-purity cultures (92.44 ± 1.66% motor neurons) co-expressing choline acetyltransferase, MNX1/HB9, and β-tubulin III, with minimal contamination from astrocytes (0.12 ± 0.01% GFAP+ cells) or microglia (0.04 ± 0.02% CD11B+ cells) [29].
Diagram 1: iPSC to Neuron Differentiation Workflow
Functional Characterization of Neuronal Cultures
Comprehensive functional characterization ensures that iPSC-derived neurons exhibit properties relevant to disease modeling and drug screening. Electrophysiological assessment is critical for confirming neuronal functionality:
Multi-electrode array (MEA) recordings of commercial iCell neurons show spontaneous, spike-like activity within 2 days of plating, peaking at one week, then decreasing to remain at low levels up to one month. These spikes are reversibly inhibited by tetrodotoxin, confirming their dependence on voltage-gated sodium channels [51].
Patch-clamp experiments demonstrate that iPSC-derived neurons generate spontaneous action potentials and express voltage-gated Na+ and K+ channels with membrane properties consistent with native neurons. The reduction in MEA spiking after the first week correlates with the development of inhibitory GABAergic tone, indicating circuit maturation [51].
Neurotransmitter receptor expression profiles confirm the presence of functional GABAₐ receptors (responsive to bicuculline, picrotoxin, and positive allosteric modulators like diazepam), glycine receptors (strychnine-sensitive), and ionotropic glutamate receptors (AMPA, kainate, and NMDA types with appropriate pharmacological properties) [51].
High-Throughput Screening Platforms and Assays
Phenotypic Screening Assays
High-content phenotypic screening in iPSC-derived neurons focuses on quantifying disease-relevant pathologies and their modulation by therapeutic compounds:
Neuronal Survival Assays implemented in the SALS motor neuron model demonstrated reduced survival of patient-derived neurons compared to controls, recapitulating a key pathological hallmark of neurodegeneration. This assay used longitudinal live-cell imaging with automated quantification to track motor neuron health over time [29].
Neurite Degeneration Metrics in the same model showed accelerated neurite degeneration in SALS motor neurons that correlated with donor survival, providing a clinically relevant phenotypic endpoint. The screening pipeline employed virally delivered motor neuron-specific reporters (HB9-turbo) for selective visualization and quantification [29].
High-Content Imaging with Machine Learning classification, adapted from malaria research, enables automated differentiation and quantification of cellular phenotypes. This approach can enumerate subcellular structures and classify morphological changes without labor-intensive manual counting [52]. For AD research, similar approaches could quantify tau pathology, synaptic density, or neurite outgrowth.
Table 2: Phenotypic Assays for Alzheimer's Disease Drug Screening
| Assay Type | Measured Parameters | Detection Method | AD Relevance |
|---|---|---|---|
| Neuronal Survival | Cell viability, apoptosis markers, caspase activation | Live-cell imaging, fluorescence microscopy | Neuronal loss in AD |
| Neurite Morphology | Neurite length, branching complexity, varicosities | High-content imaging, automated analysis | Early synaptic pathology |
| Protein Aggregation | Aβ plaque load, tau tangles, TDP-43 pathology | Immunofluorescence, FRET-based sensors | Core AD pathology |
| Synaptic Function | Synaptic density, spine morphology, synaptic markers | Electron microscopy, super-resolution imaging | Synaptic loss correlates with cognitive decline |
| Network Activity | Synchronous bursting, spike rate, oscillation patterns | Multi-electrode arrays, calcium imaging | Network hyperexcitability in AD |
Compound Screening Strategies
Systematic compound screening in iPSC-derived neuronal libraries enables identification of candidate therapeutics:
Clinical Trial Compound Retesting in the SALS model screened drugs previously tested in ALS clinical trials and found that 97% failed to mitigate neurodegeneration, reflecting actual trial outcomes and validating the model's predictive value [29].
Combinatorial Therapy Testing identified baricitinib, memantine, and riluzole as a promising therapeutic combination for sporadic ALS, demonstrating enhanced efficacy compared to individual compounds [29]. This approach is particularly relevant for AD, where multi-target therapies may address the complex, multifactorial pathology.
Polygenic Risk-Stratified Screening using resources like IPMAR enables identification of therapeutics that may be specifically effective for particular genetic subtypes of AD, moving toward personalized medicine approaches [30].
Experimental Protocols for Key Assays
Motor Neuron Differentiation and Screening Protocol
The optimized protocol for motor neuron differentiation and phenotypic screening consists of five stages [29]:
Neural Induction: Dual-SMAD inhibition using small molecule inhibitors (SB431542 and LDN193189) for 7-10 days to specify neural ectoderm fate.
Anterior-Posterior Patterning: Treatment with retinoic acid (0.1 µM) and purmorphamine (Sonic Hedgehog agonist, 1 µM) for 7 days to generate spinal motor neuron progenitors.
Motor Neuron Differentiation: Withdrawal of mitogens and addition of neurotrophic factors (BDNF, GDNF, CNTF at 10 ng/mL each) to promote terminal differentiation over 14 days.
Maturation: Extended culture in neurotrophic factors with cAMP analogs (dibutyryl cAMP, 1 mM) for 21-28 days to enhance functional maturity.
Phenotypic Screening: Plating in 384-well plates for high-content imaging, treatment with compounds, and longitudinal tracking of neuronal health using live-cell imaging with motor neuron-specific reporters.
High-Content Imaging and Analysis Protocol
A generalized protocol for high-content imaging and analysis of iPSC-derived neurons [52]:
Sample Preparation: Plate iPSC-derived neurons in matrigel-coated 384-well imaging plates at optimized density (20,000-50,000 cells/well). Allow attachment for 24 hours before compound treatment.
Staining Protocol: Fix with 4% paraformaldehyde for 15 minutes, permeabilize with 0.1% Triton X-100 for 10 minutes, block with 5% BSA for 1 hour. Incubate with primary antibodies (βIII-tubulin, MAP2, Tau, Synapsin) overnight at 4°C, followed by species-appropriate fluorescent secondary antibodies for 2 hours at room temperature. Counterstain with Hoechst 33342 for nuclei visualization.
Image Acquisition: Acquire 20-30 fields per well using a high-content imaging system (e.g., ImageXpress Micro Confocal or Opera Phenix) with 20x or 40x objectives. Maintain consistent exposure settings across plates.
Machine Learning-Based Analysis: Train classifiers using morphological features to automatically identify and quantify:
- Neuronal soma count and viability
- Neurite length and branching complexity
- Synaptic puncta density and distribution
- Pathological protein aggregation
Quality Control Metrics: Include Z' factor calculations for each assay plate (>0.5 acceptable), positive and negative controls on every plate, and normalization to plate-level controls to minimize batch effects.
The Scientist's Toolkit: Essential Research Reagents
Table 3: Essential Research Reagents for iPSC-Based Neuronal Screening
| Reagent Category | Specific Examples | Function | Application in AD Research |
|---|---|---|---|
| Reprogramming Factors | OCT3/4, SOX2, c-MYC, KLF4 (Yamanaka factors) | Somatic cell reprogramming to pluripotency | Generating patient-specific iPSC lines |
| Neural Induction Agents | SB431542 (TGFβ inhibitor), LDN193189 (BMP inhibitor) | Dual-SMAD inhibition for neural specification | Directing differentiation toward cortical neurons |
| Neuronal Maturation Factors | BDNF, GDNF, NGF, NT-3 (10-20 ng/mL) | Enhance neuronal survival, maturation, synaptic development | Promoting functional maturity relevant to AD phenotypes |
| Neuronal Markers | βIII-tubulin, MAP2, NeuN, Synapsin, PSD-95 | Identification and quantification of neuronal populations | Assessing synaptic integrity in AD models |
| AD Pathological Markers | Anti-Aβ, phospho-Tau, TDP-43 antibodies | Detection of protein aggregates and pathological hallmarks | Quantifying disease-relevant pathology |
| Cell Viability Indicators Calcein-AM, propidium iodide, caspase assays | Assessment of cell health and death mechanisms | Measuring neuroprotective effects of compounds | |
| Functional Probes | Fluo-4 AM (calcium), FM dyes (synaptic vesicle recycling) | Monitoring neuronal activity and synaptic function | Assessing functional rescue of network abnormalities |
Data Analysis and Visualization Approaches
Quantitative Analysis Frameworks
Robust data analysis is essential for extracting meaningful insights from high-throughput screening data:
Multi-Parametric Phenotypic Profiling combines multiple readouts (neuronal survival, neurite integrity, synaptic density) into composite scores that better capture therapeutic effects than single parameters alone.
Polygenic Risk Stratification analyzes compound responses according to genetic background, enabling identification of subtype-specific therapeutics. The IPMAR resource demonstrates how PRS can be used to stratify patient-derived lines for more targeted screening [30].
Longitudinal Data Modeling uses mixed-effects models to analyze temporal patterns in neuronal degeneration and treatment effects, accounting for donor-to-donor variability and time-dependent compound effects.
Visualization Strategies for Complex Screening Data
Effective visualization communicates complex screening results clearly:
Heatmaps display compound efficacy across multiple parameters and donor lines, revealing patterns of selectivity and potential subtype-specific effects [53].
Scatter Plots compare expression changes or phenotypic responses between conditions, highlighting consistently effective compounds across genetic backgrounds [53].
Dumbbell Plots visualize treatment effects by showing pre- and post-treatment values connected by lines, effectively communicating compound efficacy across multiple cellular endpoints [54].
Diagram 2: Screening Data Analysis Workflow
High-throughput and high-content screening in iPSC-derived neuronal libraries represents a transformative approach for Alzheimer's disease drug discovery. The integration of genetically diverse patient-derived models, sophisticated differentiation protocols, and multiparametric phenotypic screening creates a powerful platform for identifying therapeutics with greater clinical predictive validity. The resources and methodologies described herein provide a framework for implementing these approaches to advance AD research.
Future developments will likely focus on increasing physiological complexity through co-culture systems incorporating glial cells, implementing 3D organoid models that better recapitulate tissue-level organization, and incorporating aging-related phenotypes through genetic manipulation or prolonged culture. Additionally, the integration of multi-omics data with high-content phenotypic screening will enable deeper understanding of compound mechanisms and facilitate the development of personalized medicine approaches for Alzheimer's disease.
The development of novel therapeutics for complex neurodegenerative diseases like Alzheimer's disease (AD) faces substantial challenges, with a failure rate exceeding 99% in clinical trials [55] [56]. In this context, drug repositioning—identifying new therapeutic uses for existing drugs—has emerged as a promising strategy to accelerate treatment development. This approach leverages existing safety, pharmacokinetic, and manufacturing data, potentially reducing development time and costs [57]. Furthermore, the multifactorial nature of AD, involving amyloid pathology, tau tangles, neuroinflammation, and synaptic dysfunction, suggests that combinatorial therapies targeting multiple pathways simultaneously may yield greater efficacy than single-target approaches [58].
The integration of induced pluripotent stem cell (iPSC) technology has revolutionized neurodegenerative disease modeling. Patient-derived iPSC models recapitulate key pathological hallmarks of sporadic diseases, providing a humanized, pathophysiologically relevant system for drug screening and validation [29]. This technical guide examines successful case studies of drug repositioning and combinatorial therapy identification, with a specific focus on mechanistic insights derived from iPSC-based models of Alzheimer's disease.
Computational Frameworks for Drug Repositioning
Advanced computational strategies now enable systematic identification of repurposing candidates by integrating multi-omics data with comprehensive drug-target networks.
Network Medicine and Systems Pharmacology
Network medicine provides a conceptual framework for understanding diseases like AD as perturbations of the human interactome rather than isolated molecular defects [55] [56]. In this paradigm, interacting genes and proteins form disease modules whose collective dysfunction underlies pathology. Systems pharmacology complements this approach by examining drug actions on entire biological networks rather than single targets.
A recent CNS-focused network medicine framework achieved impressive performance in predicting viable repurposing candidates for AD, with machine-learning-validated BBB penetration prediction reaching 95.7% accuracy (AUC-ROC = 0.992) [55] [56]. This framework integrated multiple data dimensions:
- Multi-omics data integration: Genomics, transcriptomics, proteomics, and metabolomics from resources like the Alzheimer's Disease Sequencing Project (ADSP) and Alzheimer's Disease Neuroimaging Initiative (ADNI) [57]
- Network proximity mapping: Calculating the topological relationship between drug targets and disease modules within the human interactome
- Modality-specific assessment: Separate evaluation frameworks for small molecules, peptides, and biologics acknowledging distinct BBB penetration mechanisms
Table 1: Top computationally-identified repurposing candidates for Alzheimer's disease
| Candidate Drug | Original Indication | Network Medicine Score | BBB Penetration Potential | Mechanistic Rationale |
|---|---|---|---|---|
| Plerixafor | Hematopoietic stem cell mobilization | 1.170 | Favorable | Immunomodulation, potential effects on neuroinflammation |
| Trofinetide | Rett syndrome (investigational) | 1.387 | Moderate | Neuroprotective, anti-inflammatory properties |
| Memantine (Validation) | Alzheimer's disease | High | Favorable | NMDA receptor antagonism - validated by model |
| Donepezil (Validation) | Alzheimer's disease | High | Favorable | Acetylcholinesterase inhibition - validated by model |
Artificial Intelligence and Multi-Omics Data Integration
AI-guided strategies are increasingly employed to analyze the rapidly growing amount of biomedical data in AD research, including clinical trials, biomarkers, multi-omics data, and real-world patient data [58]. These approaches leverage:
- Genetic and genomic data: Genome-wide association studies (GWAS) have identified approximately 100 AD susceptibility loci [57]
- Transcriptomic signature reversal: Identifying compounds that counteract disease-associated gene expression changes
- Knowledge graph approaches: Matching disease nodes and networks to known drug nodes and networks to discover repurposing potential
These computational methods successfully identified baricitinib, an arthritis medication, as a repurposing candidate for COVID-19, demonstrating the power of network-based approaches [55] [56]. Similar methodologies are now being applied to AD, with bumetanide being nominated as a repurposable agent for APOE4 carriers based on transcriptomic studies [57].
iPSC-Based Disease Modeling for Therapeutic Screening
iPSC technology enables the generation of patient-specific neuronal models that recapitulate disease pathology, providing a powerful platform for drug screening and validation.
iPSC Model Generation and Characterization
The foundational protocol for generating iPSC-derived neurons involves a multi-stage process that has been rigorously optimized for neurodegenerative disease modeling [29]:
- iPSC generation: Fibroblasts from patient skin biopsies are reprogrammed using non-integrating episomal vectors to ensure genomic integrity
- Neural induction: Sequential application of patterning factors to direct pluripotent cells toward neural lineages
- Motor neuron differentiation: Specific protocol adaptations for spinal motor neuron generation using established developmental cues
- Neuronal maturation: Extended culture period with optimized conditions to achieve electrophysiologically functional neurons
Quality control measures include:
- Confirmation of genomic integrity, pluripotency, and trilineage differentiation potential
- Immunocytochemical validation of cell-type specific markers (e.g., ChAT, MNX1/HB9, Tuj1 for motor neurons)
- Assessment of culture purity, with optimized protocols achieving >92% motor neuron populations [29]
Diagram 1: iPSC-based modeling workflow for drug screening.
Phenotypic Screening in Sporadic Disease Models
A landmark study demonstrating the power of iPSC models for sporadic neurodegenerative disease involved the generation of an iPSC library from 100 sporadic ALS (SALS) patients [29]. This approach addressed a critical gap in the field, as most previous models focused on familial forms with known mutations.
Key phenotypic findings in SALS motor neurons included:
- Reduced survival compared to healthy controls
- Accelerated neurite degeneration that correlated with donor survival
- Transcriptional dysregulation consistent with postmortem spinal cord tissues from ALS patients
- Pharmacological rescue by riluzole, validating the model's clinical predictive value
This large-scale model demonstrated exceptional predictive validity when screening drugs previously tested in ALS clinical trials: 97% failed to mitigate neurodegeneration, accurately reflecting clinical trial outcomes [29].
Case Studies in Drug Repositioning and Combinatorial Therapy
Case Study 1: Repositioning of Baricitinib, Memantine, and Riluzole for ALS
The iPSC-based screening of SALS models identified a promising combinatorial therapy consisting of baricitinib, memantine, and riluzole [29].
Baricitinib is a JAK-STAT inhibitor approved for rheumatoid arthritis that reduces neuroinflammatory signaling. Memantine is an NMDA receptor antagonist approved for Alzheimer's disease that reduces excitotoxicity. Riluzole is the only approved medication for ALS that extends life, modulating glutamate signaling.
The experimental protocol for identifying this combination involved:
- Large-scale drug screening: Testing >100 drugs previously evaluated in ALS clinical trials
- Efficacy assessment: Only 3% of tested drugs rescued motor neuron survival across SALS donors
- Combinatorial testing: Systematic evaluation of effective drugs in combination
- Validation: The triple combination significantly increased SALS motor neuron survival across diverse patient-derived lines
This finding represents the first therapeutic candidates identified and validated across SALS donors to encompass heterogeneity in drug efficacy within the sporadic patient population [29].
Case Study 2: Network-Medicine Guided Discovery for Alzheimer's Disease
A comprehensive CNS-focused network medicine framework identified several promising repurposing candidates for AD [55] [56]. The methodological workflow included:
- Target identification: Multi-dimensional network pharmacology with temporal dynamics applied to 742 robustly dysregulated genes, generating 25 high-priority targets (e.g., IGF1, SNCA, SOX9)
- Compound filtering: CNS-specific pre-filtering of 24,474 DGIdb compounds to 8,247 CNS-relevant drugs
- Medicinal chemistry assessment: Molecular property analysis, BBB penetration prediction, chemical tractability classification, and safety evaluation
- Candidate prioritization: Modality-specific ranking of small molecules, peptides, and biologics
This approach identified plerixafor (a CXCR4 antagonist used for stem cell mobilization) as the top-ranked small molecule, and trofinetide (investigational for Rett syndrome) as the top-ranked peptide [55] [56]. The framework successfully identified FDA-approved AD therapeutics memantine and donepezil among top candidates, validating its computational performance.
Table 2: Key research reagents for iPSC-based neurodegenerative disease modeling
| Research Reagent | Function | Application in Featured Studies |
|---|---|---|
| Non-integrating episomal vectors | Reprogramming without genomic integration | Generating footprint-free iPSCs from patient fibroblasts [29] |
| HB9-turbo fluorescent reporter | Motor neuron-specific labeling | Live-cell imaging and quantification of motor neuron health [29] |
| ChAT, MNX1/HB9, Tuj1 antibodies | Cell-type identification and purity assessment | Immunocytochemical validation of motor neuron differentiation [29] |
| Multi-electrode arrays (MEAs) | Functional neuronal activity assessment | Electrophysiological characterization of neuronal networks [29] |
| RNA sequencing platforms | Transcriptomic profiling | Identification of disease-associated gene expression changes [29] |
| Live-cell imaging systems | Longitudinal tracking of neuronal health | Monitoring neurite degeneration and cell survival [29] |
Alzheimer's Disease Clinical Trial Landscape for Combinatorial Therapies
As of January 2025, 30 pharmacological drug combinations from 53 interventional clinical trials registered on ClinicalTrials.gov have been evaluated since 2015 [58]. Sixteen of these combinations have failed in completed, terminated, or withdrawn trials, highlighting the challenges in combination therapy development.
Promising combinations currently in ongoing trials include:
- Ciprofloxacin + Celecoxib (PrimeC): Targeting neuroinflammation
- Dasatinib + Quercetin (D+Q): Senolytic combination targeting cellular senescence
- E2814 + Lecanemab: Anti-tau + anti-amyloid combination therapy
- Dextromethorphan + Bupropion (AXS-05): Targeting neuropsychiatric symptoms
Diagram 2: Combinatorial therapies targeting multiple Alzheimer's pathways.
Experimental Protocols and Methodologies
Protocol for Large-Scale Drug Screening in iPSC-Derived Neurons
The following detailed protocol is adapted from the landmark SALS screening study [29]:
Stage 1: iPSC-derived motor neuron differentiation
- Utilize a rigorously optimized 5-stage protocol adapted from established spinal motor neuron differentiation methods
- Culture cells in 384-well plates suitable for high-throughput screening
- Confirm differentiation efficiency (>92% motor neurons) via immunocytochemistry for ChAT, MNX1/HB9, and Tuj1 at day 35-42
Stage 2: Longitudinal live-cell imaging and phenotypic assessment
- Transduce cultures with HB9-turbo fluorescent reporter for motor neuron-specific labeling
- Implement automated imaging systems for daily monitoring of neuronal health
- Quantify key parameters: cell survival, neurite length, branching complexity, and somal size
- Establish baseline degeneration rates in control vs. disease lines before compound addition
Stage 3: Compound library screening
- Prepare compound libraries at appropriate concentrations (typically 1-10 μM for small molecules)
- Include positive controls (e.g., riluzole for ALS models) and vehicle controls
- Add compounds to cultures at day 35 of differentiation
- Refresh compound-containing media every 72 hours
Stage 4: Combinatorial testing
- Test effective single agents in systematic combinations
- Use statistical modeling to identify synergistic, additive, or antagonistic interactions
- Validate promising combinations across multiple patient-derived lines to account for disease heterogeneity
Stage 5: Transcriptomic and functional validation
- Perform RNA sequencing on treated vs. untreated cultures to identify pathway-level effects
- Conduct electrophysiological assessments using multi-electrode arrays (MEAs) for functional validation
- Analyze correlation between phenotypic rescue and molecular signatures
Protocol for Network Medicine-Based Candidate Prioritization
The computational framework for identifying repurposing candidates involves [55] [56]:
Stage 1: Data collection and preprocessing
- Curate Alzheimer's disease gene expression datasets from GEO (e.g., GSE48350, GSE5281)
- Implement rigorous quality control: remove genes with >50% missing values, filter low-variance genes (bottom 10th percentile)
- Apply batch correction methods where appropriate
Stage 2: Disease module identification
- Identify robustly dysregulated genes through differential expression analysis
- Construct disease-specific networks using protein-protein interaction databases
- Calculate network centrality measures to identify key regulatory nodes
Stage 3: Compound-disease network integration
- Map drug-target interactions from DGIdb and other compound databases
- Calculate network proximity between drug targets and disease modules
- Generate multi-dimensional scores integrating network proximity, expression changes, and genetic evidence
Stage 4: CNS-focused filtering
- Apply blood-brain barrier penetration predictors with machine learning approaches
- Assess chemical tractability for CNS targets
- Evaluate safety profiles based on known clinical use
Stage 5: Candidate prioritization and validation
- Generate modality-specific rankings (small molecules, peptides, biologics)
- Classify evidence levels from mechanistic to clinical
- Validate predictions using independent datasets and experimental models
The integration of advanced computational approaches with human iPSC-based disease models creates a powerful framework for identifying and validating repurposed drugs and combinatorial therapies for Alzheimer's disease and related neurodegenerative conditions. The case studies of baricitinib, memantine, and riluzole in sporadic ALS models demonstrate how this integrated approach can identify promising combinations that target multiple disease mechanisms simultaneously.
The future of neurodegenerative disease therapy development will likely involve:
- Increased use of patient-derived iPSC models that capture the heterogeneity of sporadic diseases
- Advanced computational methods that integrate multi-omics data with clinical information
- Focus on combinatorial approaches that address the multifactorial nature of neurodegeneration
- Platform trials that efficiently evaluate multiple therapeutic candidates under a single protocol
These approaches offer hope for accelerating the development of effective treatments for Alzheimer's disease and related dementias, addressing a critical unmet medical need with growing societal impact.
Overcoming iPSC Model Limitations: Standardization, Reproducibility, and Phenotypic Robustness
Addressing Donor-to-Donor Variability and Ensuring Cohort Representation in iPSC Libraries
The use of induced pluripotent stem cell (iPSC) technology has established a new frontier in modeling human diseases, especially for neurological disorders like Alzheimer's disease (AD) where access to human brain tissue is limited and animal models show poor translational relevance [59] [34]. However, the thousands of human iPSC lines generated over the past decade introduce substantial challenges for experimental reproducibility and meaningful data interpretation [59]. Donor-to-donor variability represents a particularly significant concern, as differences between donor individuals have been found to affect most iPSC cellular traits, from DNA methylation and mRNA/protein abundance to pluripotency, differentiation potential, and cell morphology [59]. For Alzheimer's disease research specifically, this variability is compounded by the polygenic nature of the common disease forms, where multiple genetic variants collectively influence disease risk [30].
Without appropriate strategies to address this variability, iPSC-based models become highly vulnerable to confounding effects that obscure biological signals of interest [59]. This technical guide provides a comprehensive framework for understanding, quantifying, and mitigating donor-to-donor variability in iPSC libraries, with specific application to modeling Alzheimer's disease mechanisms. We present rigorous quality control measures, standardized protocols, robust experimental designs, and analytical approaches that enable researchers to either account for experimental variation between studies or even exploit it for deeper biological insight into AD pathogenesis.
Genetic and Non-Genetic Contributions to Variability
Systematic assessments of variability in iPSC models have revealed that genetic background constitutes the most significant source of variation. Large-scale transcriptional profiling of 317 human iPSC lines from 101 individuals demonstrated that approximately 50% of genome-wide expression variability is explained by variation across individuals [60]. Proteomic analyses similarly identified genetic background as the highest source of variability in iPSC derivatives, with inter-donor variability significantly exceeding interclonal variability [61].
Table 1: Primary Sources of Variability in iPSC Libraries
| Variability Source | Contribution Magnitude | Impacted Cellular Features | References |
|---|---|---|---|
| Genetic background (inter-individual) | 5-46% of phenotypic variation; ~50% of gene expression variation | Gene expression, eQTLs, DNA methylation, differentiation potential, morphology | [59] [60] |
| Intra-individual (interclonal) | Significantly smaller than inter-individual | Gene expression profiles, differentiation efficiency | [61] [60] |
| Technical factors (reprogramming, culture) | Contributes to residual variation after accounting for genetics | Cellular heterogeneity, maturation state, transcriptional profiles | [59] |
| Sex differences | ~2% of gene expression variation | X and Y chromosome genes | [60] |
The HipSci consortium reported that 5-46% of the variation in iPSC cell phenotypes is attributable to inter-individual differences [59]. This genetic influence persists through reprogramming and differentiation, with iPSC lines derived from the same individual being more similar to each other than to lines from different individuals at the level of gene expression, epigenetic markers, and differentiation propensity [59] [60]. This retained donor-specific signature means that iPSCs effectively maintain a genetic memory of their donor origin throughout reprogramming.
Non-genetic sources of variability include the cell type of origin, reprogramming method, culture conditions, passage number, and differentiation protocols [59]. The process of induced reprogramming involves erasing the existing epigenetic state of the somatic cell of origin, but the tissue source can still influence the propensity of a line to differentiate into different cell types [59]. Additionally, even after controlling for genotype, substantial experimental heterogeneity remains, which can confound disease modeling efforts, especially when studying small effect sizes [59].
Analytical Framework for Quantifying Variability
To systematically assess variability, researchers can employ linear mixed models that partition the total variance into contributions from different variables in the experimental design (e.g., donor, sex, reprogramming batch) plus residual variance [60]. The variancePartition method provides a statistical and visualization framework that fits a linear mixed model for each gene and quantifies the percentage of expression variance explained by each factor [60].
This approach revealed that after accounting for technical factors (sequencing batches, RNA preparation methods), variation across individuals explains a median of 49.9% of contribution to expression variance in iPSCs [60]. The residual variation (42.3% of total variance) primarily represents variation within multiple iPSC lines derived from the same individual, highlighting that both between-donor and within-donor sources must be considered in experimental design [60].
Figure 1: Sources of Variability in iPSC Libraries. Genetic factors contribute approximately 50% of the total variability in gene expression and cellular phenotypes, with non-genetic and technical factors accounting for the remainder.
Strategic Framework for Managing Variability in Alzheimer's Disease Modeling
Donor Selection and Cohort Design
For Alzheimer's disease research, appropriate donor selection is paramount. Common AD is polygenic, with heritability estimates between 60-80% for late-onset AD and 92-100% for early-onset forms [30]. The IPMAR Resource (iPSC Platform to Model Alzheimer's Disease Risk) exemplifies a strategic approach, selecting donors based on polygenic risk scores (PRS) to capture genetic extremes [30]. This resource includes 90 iPSC lines with extremes of global AD polygenic risk: 34 from high-risk late-onset AD donors, 29 from high-risk early-onset AD donors, and 27 from low-risk cognitively healthy controls [30]. An additional 19 lines capture complement pathway-specific genetic risk [30].
Table 2: Cohort Design Strategies for Alzheimer's Disease iPSC Libraries
| Strategy | Implementation | Advantages | Example |
|---|---|---|---|
| Polygenic Risk Score Stratification | Select donors from extremes of AD polygenic risk distribution | Captures polygenic architecture of common AD; enables comparison of high-risk vs low-risk genetic backgrounds | IPMAR Resource: 63 high PRS AD, 27 low PRS controls [30] |
| Isogenic Line Engineering | Gene editing (e.g., CRISPR/Cas9) to introduce or correct risk variants in controlled background | Controls for genetic background; isolates effect of specific variants | Introduction of APOE ε4 allele or other GWAS-identified variants into control lines |
| Family-Based Designs | Select related donors with differential disease status | Controls for shared genetic background; identifies modifying factors | Sibling pairs discordant for AD status |
| Pathway-Specific Stratification | Focus on genetic risk within specific biological pathways | Reveals pathway-specific mechanisms; identifies subtype-specific pathologies | Complement pathway PRS selection in IPMAR [30] |
When designing iPSC cohorts for AD research, sample size requirements must account for the polygenic nature of the disease. Existing resources that use common AD donors without selecting based on polygenic risk may not adequately capture the genetic complexity underlying the disease [30]. The substantial genetic variation between individual donors unrelated to the AD phenotype necessitates larger sample sizes to accurately distinguish differences between patient and control-derived cell lines [30].
Quality Control and Standardization Methods
Robust quality control measures are essential for managing variability. The NHLBI NextGen Consortium implemented a multi-tiered QC approach including:
- Identity confirmation: Forensic genetics comparing genome-wide genotyping data with RNA-seq data to exclude cross-contaminated lines (5.9% of lines in one analysis) [60]
- Pluripotency assessment: Principal component analysis using expression of 23 established pluripotency markers and differentiation marker NCAD to identify partially reprogrammed or differentiated lines [60]
- Chromosomal analysis: Detection of aberrations using gene expression data to exclude genetically abnormal clones [60]
For long-term cultures, automated culturing platforms can significantly reduce technical variability. The Fluent automation workstation enables systematic, reproducible liquid-handling steps (cell plating, media changes, treatment, fixation) and maintains consistent neuronal cultures for up to 6 months [62]. This approach achieves robust assay performance with z-factors ranging from 0.5-0.7, indicating excellent assay reliability for high-throughput applications [62].
Standardized differentiation protocols are equally critical. For neural models of AD, established methods include dual SMAD inhibition using Noggin and SB431542 to generate PAX6+ neural cells competent of rosette formation within 11 days [23]. Inducible systems expressing neurogenic transcription factors (NGN2, ASCL1) can produce homogeneous iPSC-derived cortical neurons within 7 days [62]. These standardized approaches reduce protocol-based variability that could otherwise confound disease-related phenotypes.
Experimental Workflows for Robust Alzheimer's Disease Modeling
Comprehensive iPSC Library Establishment and Validation
Figure 2: Experimental Workflow for Establishing Variability-Minimized iPSC Libraries for Alzheimer's Disease Research. This comprehensive pipeline begins with strategic donor selection and progresses through standardized reprogramming, differentiation, and phenotypic validation stages.
The Scientist's Toolkit: Essential Research Reagents and Platforms
Table 3: Research Reagent Solutions for Variability-Managed iPSC Research
| Reagent/Platform | Function | Application in AD Modeling | References |
|---|---|---|---|
| Sendai virus vectors | Non-integrating reprogramming | Footprint-free iPSC generation from AD patients and controls | [60] |
| Dual SMAD inhibition (Noggin, SB431542) | Neural induction from pluripotent state | Highly efficient generation of PAX6+ neural stem cells | [23] |
| Inducible NGN2/ASCL1 systems | Rapid, synchronous neuronal differentiation | Production of homogeneous cortical neuron populations | [62] |
| Automated culturing platforms (e.g., Fluent) | Standardized long-term cell maintenance | Consistent neuronal cultures for up to 6 months; high-content screening | [62] |
| Aβ42 oligomer preparations | Induction of AD-related pathologies | Modeling amyloid-induced synapse loss, tau phosphorylation, neurite degeneration | [62] |
| Multiplex staining panels | Simultaneous detection of multiple neuronal markers | Quantifying synapses, dendrites, axons, and AD pathologies in high-content formats | [62] |
Analytical Approaches for Accounting and Correcting Variability
Statistical and Bioinformatics Methods
Advanced statistical approaches can account for residual variability after experimental controls. Several methods have been developed specifically for iPSC data analysis:
- Removal of unwanted variation (RUV): A normalization method that identifies and removes unwanted sources of variation within omics readouts [59]
- Probabilistic estimation of expression residuals (PEER): Based on factor analysis, PEER takes transcript profiles and covariates as input and outputs hidden factors that explain much of the expression variability [59]
- VariancePartition: Linear mixed model framework that quantifies the percentage of expression variance explained by each variable in experimental design [60]
- Expression quantitative trait loci (eQTL) mapping: Identifies genetic variants associated with gene expression changes, helping distinguish genetic from non-genetic variability sources [59] [60]
These methods are particularly important for Alzheimer's disease modeling, where polygenic risk is influenced by numerous small-effect variants that collectively contribute to disease pathogenesis [30]. The IPMAR resource demonstrates how PRS-based selection coupled with these analytical approaches can enhance detection of disease-relevant signals [30].
Experimental Design Considerations
The proteomic assessment of variability in iPSC derivatives revealed that including more donors has a much larger influence on statistical power than adding more clones per donor [61]. This finding has profound implications for experimental design in AD research:
- Prioritize donor number over clones per donor: For fixed resources, maximizing the number of distinct donors provides greater statistical power than generating multiple clones from fewer donors [61]
- Balance case-control groups: Ensure matched distributions of age, sex, and ancestry between AD and control cohorts to minimize confounding [30]
- Incorporate repeated measures: When possible, use the same iPSC lines across multiple differentiated cell types (neurons, astrocytes, microglia) to assess cell-type-specific effects [62]
- Include reference lines: "Rosetta lines" - iPSC lines used across multiple laboratories - enable cross-study comparison and help address experimental variation between laboratories [59]
For complex phenotypic analyses in AD models, such as automated quantification of synaptic density, neurite morphology, and protein aggregation, high-content imaging with appropriate normalization controls is essential. The platform described by [62] achieves this through automated confocal image acquisition with precise segmentation of dendrites (MAP2), cell bodies (CUX2), axons (Tau, p-Tau), and synapses (Synapsin 1/2), enabling robust quantification of AD-related pathologies.
Addressing donor-to-donor variability is not merely a technical challenge in iPSC-based Alzheimer's disease research - it is a fundamental requirement for generating meaningful, reproducible insights into disease mechanisms. The strategies outlined in this technical guide, from deliberate donor selection based on polygenic risk scores to standardized differentiation protocols and advanced analytical methods, provide a comprehensive framework for managing this variability. By implementing these approaches, researchers can transform variability from a confounding factor into a source of biological insight, ultimately accelerating the development of effective therapeutics for Alzheimer's disease through more predictive human iPSC-based models.
The advent of induced pluripotent stem cell (iPSC) technology has revolutionized the study of human disease by providing in vitro access to patient-specific neurons and glial cells. This is particularly crucial for Alzheimer's disease (AD), the leading cause of dementia, where current animal models do not fully replicate human pathophysiology and effective treatments remain limited [12]. However, a significant challenge persists: conventional iPSC-derived neurons exhibit a fetal-like molecular signature and rejuvenated cellular state that fails to recapitulate the aged environment in which late-onset neurodegenerative diseases like AD manifest [63]. Reprogramming somatic cells from aged donors to pluripotency resets age-associated markers, including telomere length, metabolic profiles, and epigenetic aging signatures, creating cells that are too "young" to exhibit key pathological features [63] [64]. This technological gap has driven the development of strategies to experimentally induce aging phenotypes in iPSC-derived neuronal lineages, enabling more accurate modeling of AD mechanisms and accelerating therapeutic discovery.
Fundamental Challenges in Modeling Aging with iPSC Systems
The Rejuvenation Effect of Cellular Reprogramming
The process of reprogramming somatic cells to pluripotency reverses cellular aging, erasing molecular and phenotypic hallmarks of aging regardless of donor age. Studies demonstrate that iPSCs generated from aged donors show restoration of telomere length, reduced oxidative stress, elimination of senescence markers, and reversion of nuclear aging features such as laminar composition [63]. Consequently, iPSC-derived neurons lack the cumulative cellular damage and functional decline characteristic of aged neurons in the human brain, presenting a fundamental limitation for modeling age-related diseases [63].
Functional Immaturity of iPSC-Derived Neurons
Beyond rejuvenation, iPSC-derived cells typically exhibit developmental immaturity. Current differentiation protocols produce neurons that more closely resemble fetal rather than adult stages, with immature electrophysiological properties, synaptic networks, and metabolic profiles [63]. This immaturity persists despite protracted in vitro culture, suggesting the presence of an intrinsic timing mechanism that mirrors developmental chronology [63]. For AD research, this means that critical age-dependent processes such as amyloid-beta accumulation, tau pathology, and inflammatory responses may not fully manifest without additional intervention to induce aging.
Strategic Approaches to Induce Aging in iPSC-Derived Neuronal Models
Four primary experimental strategies have emerged to overcome the limitations of rejuvenated iPSC models and accelerate age-related phenotypes for AD research.
Progerin Overexpression
The forced expression of progerin, a truncated form of lamin A associated with Hutchinson-Gilford progeria syndrome, can induce accelerated aging phenotypes in iPSC-derived cells. When expressed in midbrain dopamine neurons, progerin triggers dendritic degeneration, inclusion body formation, and accumulation of DNA damage and mitochondrial ROS [64]. However, this approach does not consistently induce cellular senescence across all cell types, suggesting incomplete recapitulation of the aging spectrum [64].
Long-Term Culture and Spontaneous Aging
Extended in vitro culture leverages time-dependent accumulation of cellular damage to mimic natural aging. For example, iPSC-derived cardiomyocytes maintained for 120 days exhibit functional deterioration, senescence markers, increased p21 expression, and lipofuscin accumulation [64]. Similarly, prolonged culture of neuronal lineages allows gradual manifestation of age-related phenotypes, though this approach requires substantial time investment and may not fully replicate in vivo aging dynamics.
Environmental Stress Induction
Exposure to sublethal stressors mimics age-associated damage accumulation. Key approaches include:
- Oxidative Stress: Treatment with hydrogen peroxide or other ROS-inducing agents to accelerate mitochondrial dysfunction and protein oxidation [64]
- Ionizing Radiation: Gamma or proton irradiation to induce DNA damage response and cellular senescence pathways [64]
- Hypoxic Conditions: Culture under reduced oxygen tension to simulate age-related vascular dysfunction and metabolic stress [64]
Cerebral organoids subjected to hypoxia show blood-brain barrier dysfunction, increased oxidative stress, and elevated secretion of inflammatory cytokines including IL-6 and TNF-α [64].
Genetic Manipulation of Aging Pathways
Direct manipulation of genes regulating aging processes can induce premature aging phenotypes:
- Telomerase Inhibition: Shortening telomeres to trigger replicative senescence and age-associated signaling pathways [64]
- Senescence Gene Expression: Overexpression of cell cycle inhibitors such as p16INK4a or p21 to induce permanent growth arrest [64]
- APOE ε4 Expression: Introduction of the major genetic risk factor for late-onset AD to model gene-environment interactions in aging [12]
Table 1: Comparison of Aging Induction Strategies for iPSC-Derived Neuronal Models
| Strategy | Key Mechanisms | AD-Relevant Phenotypes | Timeframe | Limitations |
|---|---|---|---|---|
| Progerin Overexpression | Nuclear envelope defects, DNA damage, mitochondrial dysfunction | Dendrite degeneration, protein aggregation, oxidative stress | 2-4 weeks | Inconsistent senescence induction; non-physiological trigger |
| Long-Term Culture | Cumulative damage, telomere shortening, epigenetic drift | Lipofuscin accumulation, functional decline, senescence markers | 3-6 months | Time-intensive; variable outcomes between cell lines |
| Environmental Stress | Oxidative damage, DNA strand breaks, metabolic stress | BBB dysfunction, neuroinflammation, mitochondrial ROS | 24 hours - 2 weeks | May induce acute stress rather than chronic aging |
| Genetic Manipulation | Telomere attrition, cell cycle arrest, pathway modulation | Disease-specific pathology (e.g., Aβ with APOE4) | 2-8 weeks | Requires genetic modification; potential pleiotropic effects |
Experimental Workflows for Aging Induction
Integrated Pipeline for Accelerated Neuronal Aging
The following workflow diagram illustrates a comprehensive experimental approach combining multiple aging induction strategies:
Molecular Pathways in Neuronal Aging
Key signaling pathways involved in neuronal aging and their experimental manipulation:
The Scientist's Toolkit: Essential Reagents and Methodologies
Table 2: Research Reagent Solutions for Aging Induction in iPSC-Derived Neuronal Models
| Reagent/Category | Specific Examples | Function in Aging Induction | Application in AD Models |
|---|---|---|---|
| Reprogramming Factors | OCT3/4, SOX2, KLF4, c-MYC (Yamanaka factors); OCT3/4, SOX2, NANOG, LIN28 | Somatic cell reprogramming to pluripotency | Generate patient-specific iPSCs from familial and sporadic AD cases [65] |
| Gene Delivery Systems | Sendai virus (non-integrating), episomal plasmids, mRNA transfection | Factor delivery for reprogramming and genetic modification | Introduce AD-related mutations (e.g., APP, PSEN1) or aging modifiers (progerin) [65] |
| Differentiation Inducers | Small molecules (CHIR99021, SB431542), patterning factors (Noggin, DKK1) | Direct lineage specification toward neuronal fates | Generate cortical neurons, microglia, and astrocytes for AD modeling [65] |
| Aging Inducers | Progerin expression vectors, H₂O₂, etoposide, hydrogen peroxide | Accelerate aging phenotypes through DNA damage and stress | Induce age-related vulnerability to AD pathology [64] |
| Senescence Markers | Antibodies against p16, p21, p53; SA-β-gal assay kits | Detection and quantification of senescent cells | Validate aging induction in neuronal cultures [64] |
| AD Pathology Assays | Aβ ELISAs, phospho-tau antibodies, multiplex cytokine panels | Quantify disease-specific protein accumulation and inflammation | Measure AD-relevant phenotypes in aged neurons [12] |
Validation of Aging and Disease Phenotypes
Comprehensive characterization of experimentally aged iPSC-neuronal models should assess both general aging markers and AD-specific pathology:
- Cellular Aging Markers: SA-β-galactosidase activity, lipofuscin accumulation, telomere length analysis, nuclear morphology, and mitochondrial ROS production [64]
- Molecular Aging Signatures: Transcriptomic profiling for age-associated genes, DNA methylation clocks, and heterochromatin loss assessment [63]
- AD-Specific Pathology: Aβ40/42 secretion, phospho-tau immunoreactivity, neuritic dystrophy, and synaptic loss [12]
- Functional Assessments: Electrophysiological recording of network activity, calcium imaging for neuronal signaling, and phagocytosis assays for microglial function [12]
Advanced 3D model systems, including cerebral organoids with incorporated microglia, demonstrate particular utility for modeling the multicellular interactions central to AD pathogenesis in an aged context [12] [64].
Applications in Alzheimer's Disease Research and Drug Discovery
The development of aged iPSC-neuronal models enables several critical applications in AD research:
- Mechanistic Studies: Investigation of age-dependent vulnerability to protein misfolding, oxidative damage, and cellular resilience in patient-specific genetic backgrounds [12]
- Compound Screening: Identification of therapeutics that mitigate age-associated pathology in human neuronal systems, including senolytics, ROS scavengers, and protein homeostasis regulators [64]
- Personalized Medicine: Assessment of individual patient responses to potential therapies based on specific genetic risk profiles (e.g., APOE genotype) [12]
- Biomarker Discovery: Identification of age-exacerbated molecular signatures that correlate with disease progression for diagnostic development [12]
These applications highlight the transformative potential of combining aging induction strategies with iPSC technology to create more clinically relevant models for understanding and treating Alzheimer's disease.
Quality Control and Batch Consistency in Large-Scale iPSC Differentiation and Screening Campaigns
The use of induced pluripotent stem cell (iPSC) technology has revolutionized in vitro modeling of human diseases, offering unprecedented opportunities to study complex neurodegenerative disorders like Alzheimer's disease (AD) in a human-relevant system [34]. For AD research, iPSCs are particularly valuable as they can be differentiated into various neural cell types—including neurons, astrocytes, and microglia—that recapitulate key aspects of human neuropathology [62] [15]. However, the tremendous potential of iPSC-derived models is contingent upon solving a fundamental challenge: ensuring rigorous quality control and batch consistency throughout large-scale differentiation and screening campaigns. Variability in iPSC differentiation outcomes can obscure disease-specific phenotypes and compromise drug screening validity, making standardization protocols essential for generating physiologically relevant and reproducible models of AD mechanisms.
Critical Quality Control Metrics for iPSC-Derived Neural Cultures
Cellular Characterization and Purity Assessment
A foundational aspect of quality control involves comprehensive characterization of the cellular composition of iPSC-derived neural cultures. Immunocytochemistry and high-content imaging are standard methodologies for quantifying the presence of specific neural cell types and ensuring culture purity aligns with experimental requirements.
Table 1: Key Cell Type Markers for Quality Control in iPSC-Derived Neural Cultures
| Cell Type | Marker Proteins | Target Purity Range | Functional Assessment |
|---|---|---|---|
| Motor Neurons | ISL1, NKX6-1, HB9 (MNX1), ChAT | ~17% (mixed culture) to >90% (enriched) [39] [29] | Electrophysiology, neurite outgrowth |
| Cortical Neurons | TUBB3 (β-III Tubulin), CUX2, MAP2 | >95% [62] | Synaptic activity, glutamate response |
| Astrocytes | S100B, GFAP | Variable (context-dependent) [39] [62] | Calcium signaling, inflammatory response |
| Neural Progenitors | NESTIN, SOX1, PAX6, MUSASHI | High in expansion phases [66] | Differentiation potential, self-renewal |
Quantification of these markers reveals significant sources of variability. For instance, one large-scale study of 433 iPSC lines differentiated into motor neurons found that ALS cultures had significantly more ISL1+ motor neurons than controls (17.0% ± 0.4% versus 14.1% ± 0.8%), with the difference primarily driven by male ALS samples [39]. This highlights how both disease status and sex can influence differentiation efficiency—critical factors that must be controlled in experimental design.
Genomic and Molecular Profiling
Beyond cellular markers, quality control extends to genomic integrity and molecular profiling. Whole-genome sequencing should be implemented to identify pathogenic variants and ensure genomic stability throughout culture expansion. In AD modeling, this is particularly relevant for lines carrying familial AD mutations in APP, PSEN1, and PSEN2 genes [15]. RNA sequencing provides an additional layer of quality assessment, with principal component analysis (PCA) often used to identify batch effects and unwanted sources of variation. Research indicates that sex-driven transcriptomic signatures can completely separate male and female samples, while the percentage of S100B+ cells correlates strongly with various principal components [39]. These molecular profiling techniques help identify and account for major confounding variables before proceeding with phenotypic screening.
Standardized Differentiation Protocols for Batch Consistency
Direct Induction Methodologies
Optimized differentiation protocols are essential for minimizing variability across large-scale iPSC campaigns. For neuronal differentiation, several methodologies have been established:
- Direct Induction with Small Molecules: A 32-day direct motor neuron differentiation protocol using dual SMAD inhibition and ventral patterning has been successfully applied to hundreds of iPSC lines [39]. This method employs specific small molecule inhibitors to direct cell fate toward the desired neuronal subtype while minimizing spontaneous differentiation.
- NGN2/ASCL1-Induced Cortical Neurons: For cortical neurons relevant to AD pathology, the inducible expression of neurogenin 2 (NGN2) and achaete-scute family bHLH transcription factor 1 (ASCL1) can generate homogeneous populations of upper-layer cortical neurons within 7 days [62]. This method, when combined with cell cycle inhibition, produces cultures with over 95% expression of cortical marker CUX2.
- Neural Stem Cell (NSC) Intermediates: A scalable, cost-effective protocol for generating NSCs/early neural progenitor cells (eNPCs) without Noggin has been developed for large-scale operations [66]. This method involves forming "neurosphere-like structures" (NLSs) that can be further differentiated into predominantly VGLUT1-positive neurons, providing a renewable source of neural cells for extended screening campaigns.
Automated Culturing Platforms
Automation represents a transformative approach for enhancing batch consistency in long-term neuronal cultures. Advanced automated culturing platforms address the challenge of maintaining neuronal health and phenotype consistency over extended periods (up to 6 months), which is crucial for modeling late-onset AD pathologies [62].
Table 2: Automated Platform Components for Consistent iPSC Culture
| System Component | Function | Implementation Example |
|---|---|---|
| Liquid Handling Workstation | Precise cell plating, media changes, treatment applications | Fluent Automation Workstation (Tecan) |
| High-Content Imaging | Automated confocal image acquisition and analysis | IN Cell Analyzer 6000 (GE Healthcare), ImageXpress Micro Confocal |
| Environmental Control | Maintained temperature, CO2, and humidity during procedures | Integrated incubator modules |
| Multiplex Staining | Simultaneous labeling of multiple cellular structures | Automated staining protocols for dendrites (MAP2), synapses (Synapsin), axons (Tau) |
This automated approach enables systematic, reproducible handling of neurons with demonstrated robustness, achieving average z-factors of 0.5-0.7 across multiple batches and experiments—indicating excellent assay reliability for high-throughput screening [62].
Technical Controls and Experimental Design
Batch Control Strategies
Implementing appropriate technical controls is essential for distinguishing true biological signals from technical variability in large-scale campaigns:
- Batch Differentiation Controls (BDCs): The same control iPSC line differentiated alongside each batch of patient lines to monitor differentiation consistency across time [39].
- Batch Technical Controls (BTCs): A control line differentiated in bulk, aliquoted, and included with each RNA-seq run as a technical reference to control for processing variability [39].
- Longitudinal Live-Cell Imaging Pipelines: Continuous monitoring of neuronal health using motor neuron-specific reporters (e.g., HB9-turbo) to track degeneration kinetics without fixed-timepoint artifacts [29].
Several specific sources of variability must be actively managed:
- Reprogramming Method Effects: The original cell source for reprogramming (T-cell vs. non-T cell PBMCs) can impart distinct gene expression signatures that persist through differentiation [39]. Establishing isogenic lines or controlling for reprogramming method in experimental design is crucial.
- Sex-Specific Differences: As identified in large-scale studies, sex drives significant transcriptomic variation that can confound disease signatures [39]. Stratification by sex during analysis or balanced experimental designs are necessary controls.
- Cell Composition Effects: The percentage of non-neuronal cells (particularly S100B+ astrocyte-like cells) strongly correlates with transcriptional variation [39]. Standardizing differentiation efficiency or including cell composition as a covariate in analyses mitigates this confounder.
Applications in Alzheimer's Disease Modeling and Drug Screening
Recapitulating AD Pathology
When properly controlled, iPSC-derived neural models can recapitulate key AD pathologies in a reproducible manner. These include:
- Aβ Plaque Formation: Co-culture systems with iPSC-derived neurons and microglia demonstrate robust Aβ plaque formation with surrounding phospho-Tau positive dystrophic neurites [62].
- Tau Pathology: Treatment with Aβ42 oligomers induces hyperphosphorylation at multiple Tau sites (S396/404, S217, S235, S400/T403/S404, and T181) observed in human AD [62].
- Neuronal Degeneration: Accelerated neurite degeneration and reduced neuronal survival have been demonstrated in iPSC-derived models, with degeneration kinetics correlating with donor survival time [29].
High-Throughput Drug Screening
Quality-controlled iPSC models enable more predictive drug screening. In one large-scale study of 100 sporadic ALS patients, less than 5% of drugs previously tested in ALS clinical trials showed efficacy in rescuing motor neuron survival, reflecting the clinical failure rate of these compounds [29]. This validation of predictive capacity underscores the importance of robust models for preclinical testing. For AD, similar approaches have identified known kinase pathways (GSK3, Fyn, DLK) as modulators of Aβ-induced toxicity, confirming the utility of these systems for target discovery [62].
Essential Research Reagents and Tools
Table 3: Research Reagent Solutions for Quality-Controlled iPSC Differentiation
| Reagent/Category | Specific Examples | Function in Differentiation/QC |
|---|---|---|
| Neural Induction Media | NP Selection Medium, NP Expansion Medium [66] | Directs pluripotent cells toward neural fate |
| Small Molecule Inhibitors | Dual SMAD inhibitors, PD0332991 (cell cycle inhibitor) [62] | Enhances synchronization and homogeneity of differentiation |
| Growth Factors | BDNF, GDNF [66] | Supports neuronal maturation and survival |
| Extracellular Matrix | Matrigel [66] | Provides substrate for neural attachment and growth |
| Cell Type Markers | ISL1, NKX6-1, TUBB3, S100B, GFAP [39] [29] | Quality control assessment of differentiation efficiency |
| Functional Assay Kits | FLIPR assays, electrophysiology systems | Validates functional maturity of derived neurons |
Establishing robust quality control and ensuring batch consistency are prerequisite conditions for generating physiologically relevant iPSC models of Alzheimer's disease. The integration of standardized differentiation protocols, automated culturing platforms, comprehensive cellular characterization, and appropriate experimental controls creates a foundation for reproducible large-scale screening campaigns. As these methodologies continue to evolve, they promise to enhance our understanding of AD mechanisms and accelerate the development of effective therapeutic interventions for this devastating neurodegenerative disorder.
Visualizations
Diagram 1: Quality Control Workflow for iPSC Neural Differentiation
Diagram 2: Automated Culturing Platform
Integrating CRISPR/Cas9 Gene Editing for Isogenic Controls and Functional Validation of Genetic Risk Factors
Alzheimer's disease (AD) research has been hampered by limited treatment options and the profound complexity of its pathogenesis, which involves multiple genetic risk factors and cellular processes [20] [67]. While induced pluripotent stem cell (iPSC) technology has revolutionized neurodegenerative disease modeling by enabling the generation of patient-specific neurons and glia, the inherent genetic variability between human donors introduces significant confounding factors in experimental studies [12] [18]. The integration of CRISPR/Cas9 genome editing with iPSC models addresses this critical challenge by enabling the creation of isogenic controls—genetically identical cell lines that differ only at specific pathogenic loci [68]. This powerful combination provides researchers with an unprecedentedly precise system for dissecting the functional consequences of genetic risk factors in AD, advancing both our mechanistic understanding and therapeutic development capabilities for this devastating disorder [20] [69].
Scientific Rationale: Addressing Key Mechanistic Questions in Alzheimer's Genetics
The convergence of CRISPR and iPSC technologies provides a powerful platform to address fundamental questions in Alzheimer's disease research [69]. Genome-wide association studies (GWAS) have identified numerous genetic variants associated with AD risk, but the functional characterization of these variants has lagged significantly behind their discovery [69] [67]. Key questions include: which specific genes are affected by non-coding risk variants; which cellular processes are perturbed; whether mutations cause loss-of-function or gain-of-toxic-function effects; through which cell types these variants exert their effects; and what factors determine the selective vulnerability of specific neuronal populations [69]. The integration of CRISPR/Cas9 with iPSC models enables systematic investigation of these questions through precise genetic manipulation in human cells that recapitulate relevant aspects of brain physiology [20] [18].
Applications in Alzheimer's Disease Modeling
CRISPR/Cas9 and iPSC integration enables several critical applications in AD research:
- Functional Validation of GWAS Findings: Precise introduction of risk-associated single nucleotide polymorphisms (SNPs) into control iPSCs allows direct assessment of their pathological consequences [68].
- Elucidation of Disease Mechanisms: Isogenic pairs facilitate clean comparisons of molecular and cellular phenotypes without genetic background noise [68] [18].
- Drug Screening and Target Discovery: Engineered lines provide physiologically relevant platforms for compound screening and therapeutic target identification [67] [9].
- Study of Cell-Type-Specific Vulnerability: Differentiation into various neuronal subtypes and glia enables investigation of cell-type-specific effects of genetic variants [69] [18].
Technical Implementation: A High-Efficiency Genome Editing Workflow for iPSCs
Recent methodological advances have dramatically improved the efficiency of precise genome editing in iPSCs. The protocol below, adapted from a high-efficiency precision genome editing method published in Scientific Reports, achieves homologous recombination rates exceeding 90% through optimization of cell survival and DNA repair pathways [68].
Optimized Workflow for Generating Isogenic iPSC Lines
Table 1: Key Steps in High-Efficiency Genome Editing of iPSCs
| Step | Procedure | Purpose | Key Reagents |
|---|---|---|---|
| 1. gRNA Design | Design guide RNA with cleavage site <10 nt from target SNP; incorporate silent PAM-disrupting mutation in repair template | Maximizes HDR efficiency; prevents re-cleavage of edited alleles | Alt-R CRISPR-Cas9 gRNA [68] |
| 2. RNP Complex Formation | Combine 0.6 µM gRNA with 0.85 µg/µL HiFi Cas9 nuclease; incubate 20-30 min at RT | Forms ribonucleoprotein complex for precise genome editing | Alt-R S.p. HiFi Cas9 Nuclease V3 [68] |
| 3. iPSC Preparation | Culture iPSCs to 80-90% confluency in feeder-free conditions; change to cloning media 1h pre-nucleofection | Ensures optimal cell health and editing receptivity | StemFlex medium, Matrigel, RevitaCell, CloneR [68] |
| 4. Nucleofection | Combine RNP complex with 5 µM ssODN repair template, 50 ng/µL pCXLE-hOCT3/4-shp53-F plasmid, and 0.5 µg pmaxGFP; nucleofect | Deliver editing components while maximizing cell survival | Nucleofector system, ssODN, p53 shRNA plasmid [68] |
| 5. Cloning & Expansion | Plate in cloning media; expand surviving colonies; isolate single-cell clones | Establish purified edited cell populations | CloneR, ReLeSR, ACCUTASE [68] |
| 6. Validation | Sequence confirmation; karyotype analysis; off-target assessment | Verify precise editing and genomic integrity | ICE analysis, G-banding, whole genome sequencing [68] |
Critical Technical Considerations for Success
Several technical factors are crucial for achieving high editing efficiency:
- P53 Inhibition: Co-transfection with p53 shRNA plasmid increases homologous recombination rates approximately 11-fold by temporarily bypassing DNA damage-induced apoptosis [68].
- Pro-Survival Supplements: Adding CloneR (10%) and RevitaCell (1%) to the cloning medium improves cell survival after nucleofection by approximately 21-fold compared to base protocols [68].
- Template Design: Incorporating silent mutations in the protospacer adjacent motif (PAM) site prevents re-cleavage of successfully edited alleles, significantly improving efficiency [68].
- Cell Line Considerations: The protocol has been successfully validated across multiple iPSC lines (including PS 1.9.1 and ADRC iPSC 5.3) with consistent high efficiency, demonstrating broad applicability [68].
Research Reagent Solutions: Essential Tools for CRISPR/iPSC Research
Table 2: Key Research Reagents for CRISPR/iPSC Genome Editing
| Reagent Category | Specific Product | Function in Workflow |
|---|---|---|
| CRISPR Components | Alt-R S.p. HiFi Cas9 Nuclease V3 | High-fidelity nuclease for precise DNA cleavage with reduced off-target effects [68] |
| Alt-R CRISPR-Cas9 gRNA | Synthetic guide RNA for targeting specific genomic loci [68] | |
| Repair Templates | Single-strand Oligonucleotides (ssODNs) | Homology-directed repair templates for introducing specific point mutations [68] |
| Cell Culture Supplements | CloneR | Improves single-cell survival after nucleofection [68] |
| RevitaCell | Antioxidant supplement that enhances cell recovery after editing [68] | |
| Plasmids | pCXLE-hOCT3/4-shp53-F | Provides temporary p53 knockdown to enhance HDR efficiency [68] |
| Validation Tools | ICE Analysis Software | Quantifies genome editing efficiency from sequencing data [68] |
| G-banding Karyotyping | Detects chromosomal abnormalities in edited clones [68] |
Applications in Alzheimer's Disease Research: From Target Validation to Therapeutic Development
The CRISPR/iPSC platform has been successfully deployed to investigate multiple aspects of Alzheimer's disease pathogenesis, providing insights that would be difficult to obtain through other experimental approaches.
Key Genetic Targets in Alzheimer's Disease
Table 3: Alzheimer's Disease Genetic Targets for CRISPR/iPSC Modeling
| Gene Target | AD Association | CRISPR Application | Key Findings |
|---|---|---|---|
| APP | Familial AD mutations, Aβ production | CRISPR reduction of APP cleavage showed decreased Aβ plaques and improved cognitive function in mouse models [70] | Editing shifted APP processing toward neuroprotective sAPPa while reducing pathogenic Aβ [70] |
| APOE | Major sporadic AD risk gene (APOE4 allele) | Epigenome editing to reduce APOE4 expression without affecting protective APOE2/3 variants [70] | Successful APOE4 reduction in patient iPSC-derived brain organoids and humanized mice [70] |
| PSEN1/PSEN2 | Familial AD mutations, γ-secretase function | Creation of knockout cell lines in neuroblastoma cells to study Aβ production mechanisms [20] | Revealed how PSEN mutations alter γ-secretase processivity and increase Aβ42/Aβ40 ratio [20] |
| EIF2AK3 | Tauopathy risk gene | Introduction of GWAS-implicated SNP (rs867529) to study mechanistic links to tau pathology [68] | Established efficient editing protocol with >90% HDR efficiency for functional studies [68] |
Signaling Pathways in Alzheimer Pathogenesis
Recent research using iPSC models has identified mTOR activation as a critical pathway in AD pathogenesis, particularly in the formation of plaque-associated axonal spheroids (PAAS) [71]. Proteomic analysis of human AD postmortem tissue revealed mTOR pathway activation within these spheroids, and pharmacological inhibition of mTOR in iPSC-derived neurons ameliorated this pathology, suggesting a potential therapeutic approach [71].
The integration of CRISPR/Cas9 gene editing with iPSC technology represents a transformative approach for establishing isogenic controls and functionally validating genetic risk factors in Alzheimer's disease research. The development of highly efficient editing protocols, achieving homologous recombination rates exceeding 90%, has dramatically reduced the time and resources required to generate precisely engineered iPSC lines [68]. This technical advancement, combined with the ability to differentiate these cells into relevant neural cell types and complex organoid systems, provides an unprecedentedly powerful platform for elucidating disease mechanisms and identifying novel therapeutic targets. As these technologies continue to evolve, they hold considerable promise for advancing our understanding of Alzheimer's disease pathogenesis and accelerating the development of effective treatments for this devastating disorder.
Best Practices for Data Analysis and Interpretation in Complex, Heterogeneous Neuronal Cultures
Human induced pluripotent stem cell (iPSC) technology has revolutionized Alzheimer's disease (AD) research by enabling the generation of patient-derived neurons, glia, and organoids that retain donor-specific genetic and molecular signatures [12]. This capability is crucial for modeling complex and polygenic forms of AD, allowing researchers to capture the extremes of polygenic risk in vitro [13]. Unlike traditional rodent models, which often lack human-specific disease features and have shorter lifespans insufficient to fully develop neurodegeneration, iPSC-derived models recapitulate patient-specific pathologies, enabling the dissection of mechanistic diversity underlying AD [12]. The nervous system consists of interacting molecules, cells, and circuits across the entire body, and important functions can occur in milliseconds or minutes, or take a lifetime [72]. This complexity is amplified in heterogeneous co-culture systems where neurons, astrocytes, microglia, and other cell types form interactive networks that more faithfully represent the brain's cellular ecosystem.
The shift toward recognizing the importance of non-neuronal cells in AD pathophysiology has been driven by genetic findings. Genome-wide association studies have identified numerous late-onset AD risk genes expressed primarily in non-neuronal cells, redirecting research focus toward better understanding glial functions during neurodegeneration [73]. This recognition necessitates the development of complex in vitro models that move beyond pure neuronal cultures to incorporate the "tripartite synapse" concept, where astrocytes and microglia closely associate with synapses to form integrated functional units [73]. In this technical guide, we outline comprehensive methodologies and analytical frameworks for deriving meaningful insights from these complex, heterogeneous neuronal cultures, with particular emphasis on their application to Alzheimer's disease research.
Experimental Design for Complex Co-Culture Systems
Strategic Cell Line Selection and Culture Composition
The foundation of robust experimental outcomes in iPSC-based AD research lies in careful cell line selection and culture composition. Donor cells should be selected to capture the genetic diversity of AD, including lines with extremes of global AD polygenic risk, as exemplified by resources like the IPMAR Resource (iPSC Platform to Model Alzheimer's Disease Risk) [13]. This resource includes iPSCs with high-risk late-onset AD (34 lines), high-risk early-onset AD (29 lines), and low-risk control (27 lines), as well as those reflecting pathway-specific genetic risk such as complement pathway mutations [13].
Defining Co-Culture Ratios and Components: The composition of heterogeneous cultures should be deliberately designed to answer specific research questions. For AD research, this typically involves incorporating the major brain cell types known to be affected in disease progression. Astrocytes play critical roles in neurotransmitter recycling and metabolic functions, while microglia, as the resident innate immune cells of the brain, are involved in synaptic pruning and act as sentinels that detect and remove pathogens, dying cells, and protein aggregates such as Aβ plaques [73]. The inclusion of these cell types in co-culture systems enables research on the complex interactions that occur between brain cell types during neurodegeneration [73].
Table 1: Recommended Cell Type Ratios for Specific Research Applications
| Research Focus | Neurons | Astrocytes | Microglia | Oligodendrocytes | Rationale |
|---|---|---|---|---|---|
| Aβ and tau pathology | 70% | 20% | 10% | 0% | Enables study of neuron-astrocyte-microglia crosstalk in protein aggregation |
| Neuroinflammation | 50% | 25% | 25% | 0% | Prioritizes immune response mechanisms and cytokine signaling |
| Synaptic function | 80% | 15% | 5% | 0% | Maintains tripartite synapse integrity while minimizing confounding inflammatory factors |
| Myelination | 60% | 15% | 5% | 20% | Supports study of axonal insulation and white matter pathology |
| Blood-brain barrier | 40% | 20% | 10% | 0% (+30% endothelial) | Models neurovascular unit dysfunction in AD |
Temporal Considerations for Disease Modeling
The timing of experiments and observations must align with the anticipated pathological progression. Familial AD (FAD) models with known mutations in APP, PSEN1, or PSEN2 may demonstrate measurable phenotypes within 4-8 weeks, while sporadic AD (SAD) models with polygenic risk factors may require extended maturation periods of 12-24 weeks to develop late-onset pathology [73] [13]. APOE genotype, particularly the APOE4 allele which increases late-onset AD risk by approximately three-fold for heterozygous carriers and 15-fold for homozygous carriers, should be factored into these temporal considerations [73].
Data Acquisition and Multimodal Monitoring
Functional Activity Recording at Multiple Scales
Recent developments in neurotechnology have significantly improved the spatiotemporal resolution and throughput for monitoring brain activity in vitro. The BRAIN Initiative has emphasized the importance of producing "a dynamic picture of the functioning brain by developing and applying improved methods for large-scale monitoring of neural activity" [72]. For functional assessment of neuronal cultures, multiple complementary approaches should be employed:
High-Density Electrophysiology: Neuropixels technology, which allows high-density simultaneous recording of the activity of hundreds of neurons, has revolutionized the field of systems neuroscience and can be adapted for in vitro applications [74]. The latest version, Neuropixels NXT, offers a more compact design that increases the detail and scale at which electrophysiological neural activity can be recorded [74]. For complex co-cultures, multi-electrode arrays (MEAs) with at least 60 electrodes provide sufficient spatial sampling to detect network-level phenomena. Continuous recording sessions should span a minimum of 10 minutes at sampling rates ≥10 kHz to capture both individual action potentials and population-level oscillations.
Calcium Imaging: Genetically encoded calcium indicators (GECIs) enable monitoring of neuronal population dynamics with single-cell resolution. For cultures expressing GCamp6s or similar indicators, image at 10-20 Hz frame rate using a 10x objective for large field-of-view or 20x for detailed cellular resolution. Optogenetic manipulation during imaging allows simultaneous perturbation and observation of network dynamics, a approach aligned with BRAIN Initiative goals to "link brain activity to behavior with precise interventional tools that change neural circuit dynamics" [72].
Metabolic and Environmental Monitoring: Incorporating sensors for continuous monitoring of glucose, lactate, oxygen, and pH provides critical context for functional data. These metabolic parameters are particularly relevant in AD models given the established connection between metabolic disturbances and disease progression [73].
Endpoint Molecular and Structural Analyses
Immunocytochemistry Panels: Comprehensive staining should target cell-type-specific markers, pathological aggregates, and synaptic elements. Essential markers include: MAP2 (neurons), GFAP (astrocytes), IBA1 (microglia), Synapsin-1 (presynaptic terminals), PSD-95 (postsynaptic densities), Aβ (amyloid plaques), and phosphorylated tau (neurofibrillary tangles). High-content imaging with automated analysis enables quantification across large culture areas.
Electron Microscopy: For ultrastructural assessment of synaptic density and morphology, focused ion beam scanning electron microscopy (FIB-SEM) provides nanometer-resolution reconstruction of synaptic contacts. This approach supports the BRAIN Initiative goal to "generate circuit diagrams that vary in resolution from synapses to the whole brain" [72].
Multi-omics Sampling: Single-cell RNA sequencing captures transcriptomic heterogeneity within cultures, while proteomic analyses quantify protein expression and post-translational modifications. For AD models, particular attention should be paid to pathways involving inflammation, endocytosis, and protein degradation, which are commonly disrupted in disease [73].
Analytical Frameworks for Multidimensional Data
Functional Network Analysis
The analysis of neural circuits requires identifying and characterizing the component cells, defining their synaptic connections, observing their dynamic patterns of activity during function, and perturbing these patterns to test their significance [72]. Several key metrics should be calculated from functional recordings:
Table 2: Key Metrics for Functional Network Analysis
| Metric Category | Specific Measures | Technical Implementation | Biological Interpretation |
|---|---|---|---|
| Single-unit activity | Firing rate, burst characteristics, waveform shape | Spike sorting (Kilosort, MountainSort) | Neuronal health and excitability |
| Synchrony | Correlation coefficients, phase-locking value | Cross-correlation, coherence analysis | Functional connectivity and network integration |
| Oscillatory power | Band-specific power (theta: 4-8 Hz, beta: 13-30 Hz, gamma: 30-100 Hz) | Fourier transform, wavelet analysis | Network-level communication and information processing |
| Information theory | Mutual information, entropy | Minimum probability flow, maximum entropy models | Coding capacity and network complexity |
| Graph theory | Node degree, clustering coefficient, path length | NetworkX, Brain Connectivity Toolbox | Topological organization and efficiency |
Integrating Multimodal Data
The BRAIN Initiative has highlighted that "rigorous theory, modeling, and statistics are advancing our understanding of complex, nonlinear brain functions where human intuition fails" [72]. New kinds of data are accruing at increasing rates, mandating new methods of data analysis and interpretation [72]. To enable progress in theory and data analysis, collaborations between experimentalists and scientists from statistics, physics, mathematics, engineering, and computer science are essential [72].
Dimensionality Reduction and Clustering: Uniform Manifold Approximation and Projection (UMAP) and t-distributed Stochastic Neighbor Embedding (t-SNE) effectively visualize high-dimensional data structures. For clustering, density-based methods like HDBSCAN identify distinct functional cell types without presuming number of clusters.
Multivariate Predictive Modeling: Random forest and gradient boosting machines handle mixed data types (continuous functional metrics, categorical molecular markers) to identify features most predictive of disease phenotype. Regularized regression (elastic net) models molecular readouts from functional data.
Dynamic Systems Approaches: Hidden Markov models capture transitions between network states, while generalized linear models with Poisson noise describe spike train relationships. Dynamic causal modeling infers effective connectivity and how it is modulated by experimental conditions.
Visualization Strategies for Complex Data
Selecting Appropriate Chart Types
Effective data visualization is crucial for interpreting and communicating results from complex neuronal cultures. The choice of visualization should match the data structure and analytical question [54].
Comparison Charts: Bar and column charts are ideal for comparing values across categories, such as mean firing rates between different experimental groups [54]. For AD models, grouped bar charts effectively display metrics across multiple cell types and conditions simultaneously. Lollipop charts serve as space-efficient alternatives when comparing many categories [54].
Composition Charts: Stacked area charts show how parts of a whole change over time, such as the proportion of different neural cell types throughout culture maturation [75]. For single-timepoint composition, treemaps display hierarchical data structures, like gene expression patterns within functional pathways [75].
Relationship Charts: Scatter plots with regression lines visualize correlations between continuous variables, such as synaptic density and network synchrony. Network graphs depict functional connectivity patterns with nodes representing neurons and edges representing connection strength.
Distribution Charts: Violin plots combine box plots with kernel density estimation to show the distribution of continuous variables like inter-spike intervals across experimental groups. Histograms reveal the shape of parameter distributions, such as synaptic puncta sizes.
Creating Accessible Visualizations
Adherence to Web Content Accessibility Guidelines (WCAG) ensures visualizations are interpretable by all readers. The minimum contrast ratio for normal text should be at least 4.5:1, and for large text, at least 3:1 [76]. Non-text elements like graphical objects and user interface components must have a contrast ratio of at least 3:1 against adjacent colors [76]. The specified color palette (#4285F4, #EA4335, #FBBC05, #34A853, #FFFFFF, #F1F3F4, #202124, #5F6368) provides sufficient contrast when appropriately paired.
The Scientist's Toolkit: Essential Research Reagents and Solutions
Table 3: Essential Research Reagents for Complex Neuronal Culture Experiments
| Reagent Category | Specific Examples | Function | Application Notes |
|---|---|---|---|
| Reprogramming factors | OCT4, SOX2, KLF4, c-MYC | Somatic cell reprogramming | Generate patient-specific iPSCs; optimize delivery method to minimize genomic integration |
| Neural induction | SMAD inhibitors (dorsomorphin, SB431542) | Direct differentiation toward neural lineage | Dual SMAD inhibition yields more homogeneous neural progenitor populations |
| Cell-type specific differentiation | BDNF, GDNF, cAMP, TGF-β, IL-34 | Promote maturation of specific neural cell types | IL-34 specifically supports microglial survival and function in co-cultures |
| Pathological induction | Synthetic Aβ42, okadaic acid, oxidative stressors | Accelerate AD-relevant pathology | Concentration and timing critical to avoid acute toxicity unrelated to AD mechanisms |
| Activity monitoring | GCaMP variants, voltage-sensitive dyes | Report neuronal activity | Consider phototoxicity in long-term imaging; select indicators matched to temporal resolution needs |
| Cell labeling | CellTracker dyes, lentiviral GFP/RFP | Distinguish cell types in co-culture | Use at minimal effective concentrations to avoid altering cellular physiology |
| Fixation & permeabilization | Paraformaldehyde, Triton X-100, Tween-20 | Preserve structure while allowing antibody access | Optimization required for different antigen-antibody combinations |
| Blocking solutions | BSA, normal serum, fish skin gelatin | Reduce nonspecific antibody binding | Match serum species to secondary antibody host when possible |
Experimental Protocols for Key Assays
Multi-Electrode Array (MEA) Recordings from Heterogeneous Cultures
Materials:
- 48- or 96-well MEA plates
- Extracellular recording solution (in mM: 145 NaCl, 5 KCl, 2 CaCl2, 1 MgCl2, 10 HEPES, 10 glucose)
- Data acquisition system with environmental control (37°C, 5% CO2)
- Analysis software (Neuroexplorer, Axograph, or custom MATLAB/Python scripts)
Procedure:
- Replace culture medium with extracellular recording solution equilibrated to 37°C.
- Allow cultures to acclimate to recording chamber for 10 minutes.
- Record spontaneous activity for 10 minutes at 10 kHz sampling rate.
- Apply test compounds directly to wells during recording for pharmacological characterization.
- Include negative controls (vehicle only) and positive controls (e.g., GABA receptor antagonists).
- For AD models, specifically test responses to Aβ oligomers and modulators of tau phosphorylation.
Analysis:
- Apply high-pass filter (300 Hz) for spike detection and low-pass filter (100 Hz) for local field potentials.
- Use automated spike sorting algorithms to isolate single units.
- Calculate network burst parameters: frequency, duration, inter-burst interval, and spikes per burst.
- Compute synchrony indices and cross-correlation matrices between electrodes.
Calcium Imaging in iPSC-Derived Co-Cultures
Materials:
- Genetically encoded calcium indicator (GCaMP6f or similar)
- Live-cell imaging medium without phenol red
- Confocal or epifluorescence microscope with environmental chamber
- 20x objective for cellular resolution or 10x for network-level analysis
Procedure:
- Transduce cultures with GCaMP6f using lentiviral delivery at time of plating.
- 14-21 days post-transduction, replace medium with imaging medium.
- Image at 10-20 Hz for 5-minute epochs with minimal laser power to reduce phototoxicity.
- Include stimulus application capability for functional connectivity mapping.
- For AD models, compare calcium transient properties between isogenic lines with different AD risk profiles.
Analysis:
- Preprocess videos with motion correction and background subtraction.
- Detect regions of interest (ROIs) corresponding to individual cells.
- Extract fluorescence traces (F) and convert to ΔF/F0.
- Identify calcium events using threshold-based detection or deconvolution approaches.
- Construct functional connectivity matrices based on event co-occurrence.
Validation and Reproducibility Framework
Quality Control Metrics
Establishing rigorous quality control standards is essential for generating reproducible data from complex neuronal cultures. The adoption of open science practices is key to advancing research in neurophysiology, facilitating improved reproducibility, broader collaboration, efficient data reuse, and deeper insights into the fundamental workings of the brain [74].
Cellular Composition Verification: Regular immunocytochemical quantification of cell-type markers ensures culture composition remains within experimental parameters. Establish acceptance criteria for each cell type percentage (e.g., neurons: 65-75%, astrocytes: 15-25%, microglia: 5-15%).
Functional Benchmarks: Define expected ranges for key functional parameters in control cultures, such as mean firing rate (0.5-5 Hz), network burst frequency (0.01-0.1 Hz), and synchrony indices (0.1-0.4). Cultures falling outside these ranges may indicate technical issues requiring investigation.
Molecular Standards: Implement reference samples with known properties included in each batch of molecular analyses. For RNA sequencing, use external RNA controls Consortium (ERCC) spikes to monitor technical performance.
Data Management and Sharing
The BRAIN Initiative has identified establishing "platforms for sharing data" as a core principle, noting that public, integrated repositories for datasets and data analysis tools, with an emphasis on ready accessibility and effective central maintenance, will have immense value [72]. Similarly, the Open Data in Neurophysiology (ODIN) initiative emphasizes that while online data repositories reduce pressure for individual researchers to manage long-term data storage themselves, they require substantial and reliable resources to scale at a rate that matches demand as data generation increases [74].
Metadata Standards: Adapt the Neurodata Without Borders (NWB) standard for in vitro data, including complete experimental details: cell line origins, culture conditions, differentiation protocols, and analysis parameters.
Repository Selection: Archive data in appropriate public repositories before publication. Raw sequencing data belongs in GEO or ArrayExpress, while functional data can be stored in the Distributed Archives for Neurophysiology Data Integration (DANDI) [74].
Code Sharing: Publish analysis scripts alongside data, using version-controlled repositories like GitHub or GitLab. Containerization with Docker or Singularity ensures computational reproducibility.
Interpreting Results in the Context of Alzheimer's Disease
Relating In Vitro Phenotypes to Clinical Manifestations
When observing phenotypes in complex neuronal cultures, careful interpretation is required to establish relevance to human AD. The field is moving toward a more holistic view of AD that incorporates the functions of multiple different cell types of the brain, recognizing that while neuron dysfunction and degeneration undoubtedly underlies much of the characteristic cognitive decline, non-neuronal cells are increasingly recognized for helping to maintain proper brain function and ensuring the long-term health and survival of neurons [73].
Functional Deficits: Reduced network synchrony and altered oscillatory patterns in vitro may correspond to disrupted large-scale brain networks observed in AD patients through EEG and fMRI. Impaired homeostatic plasticity in cultures might reflect the circuit hyperexcitability and silent seizures documented in AD patients.
Molecular Pathways: Elevated Aβ42/Aβ40 ratios in culture media recapitulate the altered APP processing observed in both familial and sporadic AD [73]. Increased phosphorylation of tau at AD-relevant epitopes suggests conserved pathological mechanisms. Activation of microglia and astrocytes in culture mirrors the neuroinflammatory component of AD progression.
Distinguishing Disease-Relevant from Incidental Phenotypes
Not all phenotypic differences between AD and control cultures necessarily reflect disease mechanisms. Careful controls are essential:
Isogenic Controls: Compare phenotypes between genetically corrected lines to confirm mutation-specific effects rather than background genetic variation.
Phenocopy Experiments: Recapitulate observed phenotypes in control cultures by introducing suspected mediators (e.g., synthetic Aβ oligomers, conditioned media from AD cultures).
Rescue Studies: Reverse phenotypes through genetic or pharmacological intervention to establish causal relationships and identify potential therapeutic targets.
The analysis and interpretation of data from complex, heterogeneous neuronal cultures represents both a formidable challenge and tremendous opportunity in Alzheimer's disease research. By implementing the comprehensive framework outlined here—spanning careful experimental design, multimodal data acquisition, sophisticated analytical approaches, rigorous validation, and contextual interpretation—researchers can maximize the potential of iPSC-based models to illuminate AD mechanisms and identify novel therapeutic strategies. As these methods continue to evolve alongside advancing technologies and open science practices, they promise to progressively bridge the gap between in vitro observation and clinical reality, ultimately accelerating progress toward effective interventions for this devastating neurodegenerative disorder.
Benchmarking iPSC Models: Correlating In Vitro Findings with Animal Models and Human Clinical Outcomes
Alzheimer's disease (AD) research urgently requires models that faithfully recapitulate human pathophysiology to bridge the translational gap between preclinical findings and clinical trial success. Induced pluripotent stem cell technology has emerged as a transformative approach, generating patient-derived neural cells that model human disease with unprecedented accuracy [77] [18]. This technical guide provides a comparative analysis of iPSC-based models against traditional animal and cell line systems, framed within the context of elucidating AD mechanisms. We examine the technical specifications, experimental applications, and methodological considerations essential for researchers and drug development professionals navigating this evolving landscape.
Alzheimer's Disease Pathophysiology and Modeling Requirements
Key Pathological Mechanisms
Alzheimer's disease involves progressive neurodegeneration driven by multifaceted pathological processes. The core hallmarks include extracellular amyloid-beta plaques and intracellular neurofibrillary tangles composed of hyperphosphorylated tau protein [18]. Beyond these canonical features, AD pathogenesis involves neuroinflammation mediated by glial cells, synaptic dysfunction, metabolic disturbances, and compromised blood-brain barrier integrity [18]. Understanding these interconnected mechanisms requires models that capture human-specific pathophysiology, particularly for sporadic AD which represents over 95% of cases and involves complex genetic-risk profiles [78] [18].
Critical Modeling Capabilities
Effective AD models must replicate several critical aspects of human disease: genetic diversity (including APOE isoforms and GWAS-identified risk variants), cell-type interactions within the neurovascular unit, age-related processes, and proteinopathy progression [78] [18]. No single model system fully captures this complexity, necessitating strategic selection based on research objectives, whether for initial target validation, mechanistic investigation, or preclinical therapeutic screening.
Traditional Model Systems in AD Research
Rodent Models
Strengths and Applications
Rodent models have formed the backbone of AD research for decades, offering integrated physiological systems that capture complex brain circuitry and behavioral outputs [77] [18]. Transgenic models expressing human FAD mutations (e.g., APP, PSEN1/PSEN2) have been instrumental in establishing the amyloid cascade hypothesis and investigating Aβ toxicity mechanisms [18]. Their defined genetic backgrounds and reproducible pathology timelines facilitate standardized therapeutic testing, while their susceptibility to genetic manipulation enables causal studies [18].
Limitations and Species Disparities
Despite their utility, rodent models exhibit significant limitations. Species differences in genetics profoundly impact AD relevance; for example, rodents possess only one APOE gene variant unlike the three human isoforms (APOE2, E3, E4) that substantially modify AD risk [78]. Mouse models overexpressing FAD mutations often produce non-physiological expression patterns and fail to fully recapitulate the tau pathology and progressive neurodegeneration seen in humans [18]. Additionally, fundamental differences in immune function, metabolism, and lifespan limit their accuracy in modeling age-dependent human neurodegenerative processes [78] [18].
Table 1: Comparative Analysis of Primary Rodent Models in Alzheimer's Research
| Model Type | Key Features | Pathological Hallmarks | Major Limitations |
|---|---|---|---|
| 5xFAD Transgenic Mice | Overexpression of 5 human FAD mutations in APP & PSEN1 | Early, aggressive Aβ deposition; synaptic dysfunction; gliosis | Non-physiological APP processing; minimal tau pathology; non-human genetic background |
| 3xTG Mice | Triple mutations in APP, PSEN1, and MAPT | Aβ plaques; tau tangles; synaptic dysfunction; cognitive deficits | Artificial mutant combinations; non-human tau isoforms |
| APP Knock-In Mice | Humanized Aβ sequence with FAD mutations knocked into mouse App locus | More physiological APP expression and processing; Aβ pathology | Milder, later-onset pathology; limited neuronal loss; species-specific microenvironment |
Immortalized Cell Lines
Applications and Practical Advantages
Immortalized cell lines offer practical experimental accessibility for AD research. Microglial lines like BV2 (mouse) and HMO6 (human) provide scalable, consistent platforms for studying neuroinflammation, phagocytosis, and cytokine signaling [79]. Their rapid expansion capabilities, high homogeneity, and suitability for high-throughput screening make them valuable for initial drug candidate evaluation and mechanistic studies under controlled conditions [79] [80].
Fundamental Limitations
The immortalization process fundamentally alters cell physiology, creating significant limitations. These lines exhibit abnormal proliferation, metabolic shifts, and divergent expression profiles compared to primary cells [79]. As continuously dividing cells, they poorly model the post-mitotic state of neurons and age-related processes central to AD. Furthermore, common lines like BV2 microglia display attenuated inflammatory responses compared to primary microglia, potentially misrepresenting neuroimmune interactions [79].
Table 2: Traditional Cell Culture Models for Alzheimer's Research
| Cell Model | Origin/Generation | Key Applications in AD Research | Technical Limitations |
|---|---|---|---|
| Primary Rodent Microglia | Isolated from postnatal brain tissue via adhesion/agitation protocols | Neuroinflammation studies; Aβ phagocytosis assays; cytokine signaling | Time-consuming isolation; limited cell yield; species-specific responses |
| BV2 Microglia Cell Line | v-raf/v-myc immortalized murine microglia | High-throughput screening; mechanistic studies of neuroinflammation | Altered inflammatory responses compared to primary cells; non-human origin |
| SH-SY5Y Neuroblastoma Line | Human neuroblastoma-derived cell line | Neuronal differentiation studies; Aβ toxicity mechanisms; drug screening | Cancerous origin; limited differentiation maturity; abnormal metabolism |
iPSC-Based Models for Alzheimer's Disease
Technological Foundations
iPSC technology leverages cellular reprogramming to convert somatic cells into pluripotent stem cells capable of differentiating into any cell type. The seminal discovery by Takahashi and Yamanaka identified that forced expression of OCT4, SOX2, KLF4, and c-MYC could reprogram adult fibroblasts to a pluripotent state [78] [81]. Subsequent advances have improved safety profiles by developing non-integrating methods including Sendai vectors, episomal plasmids, and mRNA reprogramming [81]. These iPSCs can be differentiated into the major brain cell types affected in AD—neurons, astrocytes, microglia, and even cerebral organoids that recapitulate aspects of tissue-level organization [18] [82].
Key Strengths for AD Research
Human Genetic Context
iPSC models preserve the complete human genetic background of donors, enabling investigation of both familial AD mutations and sporadic AD polygenic risk factors in their native genomic context [78] [18]. This is particularly crucial for studying APOE isoforms, TREM2 variants, and other GWAS-identified risk genes that have human-specific functions not adequately modeled in rodents [18] [82]. The ability to generate isogenic lines through CRISPR/Cas9 gene editing allows precise determination of causal genetic contributions by comparing edited and parent lines while maintaining identical genetic backgrounds [78] [9].
Patient-Specific Modeling
iPSCs enable personalized disease modeling by capturing the genetic diversity of human populations [77] [83]. Researchers can generate cell lines from patients with specific genetic risk profiles, clinical presentations, or treatment responses, facilitating stratified drug development and personalized therapeutic approaches [83] [18]. This capability addresses a critical limitation of traditional models that utilize limited, standardized genetic backgrounds poorly representing human diversity.
Comprehensive Cellular Environments
Advanced differentiation protocols now yield functionally mature human neurons that develop AD-relevant pathology including Aβ accumulation and hyperphosphorylated tau [18]. iPSC-derived microglia exhibit authentic human immune functions such as Aβ phagocytosis, cytokine secretion, and synaptic pruning—capabilities that differ significantly from rodent microglia or immortalized lines [18] [82]. Similarly, iPSC-derived astrocytes recapitulate human-specific metabolic support functions and inflammatory responses [82]. The development of co-culture systems and 3D organoids enables investigation of cell-cell interactions within increasingly complex microenvironments that mimic the brain's cellular ecosystem [18].
Direct Comparative Analysis: iPSC vs. Traditional Models
Methodological and Technical Comparison
Table 3: Comprehensive Comparison of AD Model Systems Across Technical Parameters
| Parameter | iPSC-Derived Models | Traditional Cell Lines | Rodent Models |
|---|---|---|---|
| Genetic Relevance | Full human genetic background; patient-specific mutations & polygenic risk | Limited or artificial genetic background; often cancerous origin | Non-human genetics; limited transgene expression |
| Cellular Diversity | All major CNS cell types (neurons, astrocytes, microglia, etc.) | Limited to specific cell types; often monoculture | Full organismal complexity but species-specific cell properties |
| Pathological Hallmarks | Aβ & tau pathology; neuroinflammation; synaptic dysfunction | Limited proteinopathy; abnormal metabolism | Aβ plaques (some models); limited tau pathology; species-specific responses |
| Throughput & Scalability | Moderate; improving with automation | High; suitable for HTS | Low; time and resource intensive |
| Temporal Resolution | Weeks to months for phenotype development | Hours to days | Months to years for pathology |
| Technical Complexity | High; requires specialized expertise | Low to moderate; widely accessible | Moderate to high; requires animal facilities |
| Cost Considerations | High initial setup; moderate per experiment | Low | Very high (housing, maintenance) |
| Pharmacological Predictive Value | Improving for human responses; some clinical validation | Limited by transformed phenotype | Moderate; multiple failures in translation |
Applications in Drug Discovery and Development
Target Identification and Validation
iPSC models excel in target identification by enabling functional studies of novel AD risk genes identified through human GWAS in their native cellular context [18] [9]. For example, investigating TREM2 function in iPSC-derived microglia has revealed human-specific roles in Aβ clearance and inflammatory signaling that differ from mouse models [82]. Similarly, APOE4 pathogenicity can be studied in human astrocytes and neurons, revealing isoform-specific effects on lipid metabolism, Aβ aggregation, and neurotoxicity [18].
Compound Screening and Lead Optimization
iPSC-based platforms enable human-relevant compound screening at scale. Phenotypic screens using iPSC-derived neurons or glial cells expressing AD-relevant pathologies can identify compounds that modify disease phenotypes [80] [9]. The development of 3D organoid systems and high-content imaging approaches further enhances the physiological relevance and information content of these screens [83] [80]. Several candidates identified through iPSC screens, including bosutinib, ropinirole, and ezogabine, have advanced to clinical trials for neurodegenerative diseases, demonstrating the translational potential of this approach [9].
Safety and Efficacy Profiling
iPSC-derived human cells provide a relevant system for assessing compound toxicity and therapeutic indices before advancing to costly clinical trials [83]. The ability to test compounds across multiple cell types (neurons, astrocytes, microglia) enables comprehensive evaluation of cell-type-specific effects and identification of potential off-target activities [18] [80]. Furthermore, panels of iPSCs from genetically diverse donors allow preliminary assessment of population variability in drug response, supporting the development of stratified medicine approaches [83].
Experimental Protocols for iPSC-Based AD Research
iPSC Differentiation to Cortical Neurons
This protocol generates functionally mature cortical neurons from human iPSCs, suitable for modeling AD-associated neurodegeneration and toxicity studies [18]. The process requires approximately 60-90 days to achieve electrophysiologically active neurons expressing mature cortical markers.
Materials and Reagents
- Essential Reagents:
- Dual-SMAD inhibition compounds (LDN-193189, SB431542) for neural induction
- Neural maintenance media (Neurobasal/B27 supplements)
- Patterning factors (Retinoic acid, FGF-2) for cortical specification
- Maturation supplements (BDNF, GDNF, cAMP)
Procedural Steps
- Neural Induction (Days 0-10): Culture iPSCs in mTeSR medium until 70% confluent. Switch to neural induction medium containing LDN-193189 (100nM) and SB431542 (10μM). Change medium daily.
- Neural Precursor Expansion (Days 10-30): Mechanically dissociate neural rosettes and plate on poly-ornithine/laminin-coated surfaces in neural expansion medium. Passage every 7-10 days.
- Cortical Patterning (Days 30-45): Add retinoic acid (100nM) and FGF-2 (20ng/mL) to promote cortical identity. Monitor expression of FOXG1 (forebrain marker) and PAX6 (cortical progenitor marker).
- Terminal Differentiation (Days 45-90): Withdraw FGF-2 and add BDNF (20ng/mL), GDNF (10ng/mL), and cAMP (500μM) to promote neuronal maturation. Assess electrophysiological activity by patch clamping after day 60.
Microglial Differentiation Protocol
This method generates functional human microglia from iPSCs through hematopoietic progenitor intermediates, requiring approximately 45-60 days [18] [82]. The resulting cells express characteristic microglial markers (P2RY12, TMEM119, IBA1) and exhibit authentic functions including phagocytosis, cytokine secretion, and process motility.
Materials and Reagents
- Essential Reagents:
- Hematopoietic cytokines (IL-3, M-CSF, GM-CSF) for myeloid differentiation
- Microglial maturation factors (IL-34, TGF-β)
- Matrigel or poly-D-lysine for coating culture surfaces
Procedural Steps
- Mesodermal Induction (Days 0-5): Differentiate iPSCs in serum-free differentiation medium with BMP4 (50ng/mL) and VEGF (50ng/mL) to induce mesodermal commitment.
- Hematopoietic Progenitor Generation (Days 5-15): Switch to hematopoietic medium containing SCF (100ng/mL), FLT3L (100ng/mL), and TPO (50ng/mL) to generate hematopoietic progenitors.
- Myeloid Differentiation (Days 15-30): Culture hematopoietic progenitors with M-CSF (100ng/mL) and IL-3 (25ng/mL) to promote myeloid lineage commitment.
- Microglial Maturation (Days 30-45): Add IL-34 (100ng/mL) and TGF-β (10ng/mL) to induce microglial identity. Validate by flow cytometry for CD11b, CX3CR1, and P2RY12 expression.
Co-culture System for Neuroinflammation Studies
This protocol establishes co-cultures of iPSC-derived neurons, astrocytes, and microglia to model neuroimmune interactions in AD [18] [82]. The system recapitulates key aspects of neuroinflammation, including cytokine-mediated toxicity, microglial phagocytosis of synapses, and glial responses to Aβ.
Materials and Reagents
- Essential Reagents:
- Cell culture inserts (0.4μm pore) for compartmentalized co-culture
- Aβ oligomers (prepared from synthetic Aβ1-42)
- Cytokine analysis kits (for TNF-α, IL-1β, IL-6 measurement)
Procedural Steps
- Basal Layer Establishment: Plate astrocytes at 50,000 cells/cm² and culture for 7 days to establish a supportive monolayer.
- Neuronal Seeding: Seed cortical neurons at 100,000 cells/cm² onto the astrocyte bed. Culture for 21 days to establish synaptic networks.
- Microglial Addition: Add microglia at a 1:10 ratio (microglia:neurons) either directly or in transwell inserts.
- Challenge Paradigms: Treat co-cultures with Aβ oligomers (500nM) or other AD-relevant insults. Assess neuroinflammation by cytokine ELISA, neuronal death by LDH release, and synaptic density by immunostaining.
Research Reagent Solutions for iPSC-based AD Modeling
Table 4: Essential Research Reagents for iPSC-based Alzheimer's Disease Modeling
| Reagent Category | Specific Examples | Application in AD Research | Technical Considerations |
|---|---|---|---|
| Reprogramming Factors | OCT4, SOX2, KLF4, c-MYC; Sendai virus; episomal vectors | iPSC generation from patient somatic cells | Non-integrating methods preferred for clinical applications; efficiency varies by method |
| Neural Induction Supplements | LDN-193189 (BMP inhibitor); SB431542 (TGF-β inhibitor); N2/B27 supplements | Neural differentiation from iPSCs | Dual-SMAD inhibition significantly improves efficiency and purity |
| Cell-Type Specific Markers | MAP2, NeuN (neurons); GFAP, S100β (astrocytes); IBA1, P2RY12 (microglia) | Characterization of differentiated cells | Multiple markers recommended for conclusive identification |
| AD Pathology Reagents | Aβ42 oligomers; tau phosphorylation antibodies (AT8, PHF-1); γ-secretase inhibitors | Induction and detection of AD-related pathology | Aβ oligomer preparation quality critical for reproducible results |
| Functional Assay Kits Calcium imaging dyes; microelectrode arrays; phagocytosis pH-sensitive probes | Assessment of neuronal function, network activity, and microglial function | Functional validation essential beyond morphological characterization |
Current Challenges and Technical Limitations of iPSC Models
Technical and Methodological Hurdles
Despite their promise, iPSC models face significant technical challenges. Line-to-line variability arising from genetic differences between donors can complicate data interpretation and require larger sample sizes [83] [81]. The immature phenotype of iPSC-derived cells often more closely resembles fetal rather than adult human brain cells, limiting their ability to model age-related neurodegenerative processes [18] [81]. Additionally, current differentiation protocols yield limited cellular diversity that doesn't fully capture the complexity of the human brain, though organoid technologies are rapidly addressing this limitation [18].
Standardization and Scalability Issues
The field currently lacks standardized protocols across laboratories, making direct comparison of results challenging [81]. Quality control metrics for characterizing iPSC lines and their differentiated progeny remain variable, though efforts are underway to establish consensus standards [81]. Scalability for high-throughput screening, while improving, still lags behind traditional cell lines due to the lengthy differentiation timelines and technical complexity [80] [9].
Tumorigenicity and Safety Concerns
A significant safety concern for therapeutic applications is the potential for residual undifferentiated iPSCs to form teratomas [81]. The original reprogramming factors, particularly c-MYC, present oncogenic risks that must be carefully managed through improved reprogramming methods and rigorous purification of differentiated cells [81]. Additionally, genomic instability acquired during reprogramming or prolonged culture necessitates comprehensive characterization before experimental or clinical use [81].
Future Directions and Concluding Perspectives
Emerging Technological Innovations
The iPSC field is advancing rapidly through several technological frontiers. Brain organoid systems are becoming increasingly sophisticated, incorporating multiple brain regions, vascularization, and improved maturation [18]. Microfluidic platforms and organs-on-chips enable precise control of microenvironmental conditions and high-resolution monitoring of cellular interactions [83]. The integration of artificial intelligence with high-content screening data is enhancing phenotypic characterization and predictive modeling [9]. Single-cell multi-omics approaches provide unprecedented resolution of cellular heterogeneity and molecular changes during disease progression.
Path Toward Clinical Translation
For iPSC models to fully realize their potential in AD drug discovery, several developments are critical: establishment of standardized validation frameworks across laboratories, creation of comprehensive iPSC biobanks representing diverse genetic backgrounds and clinical subtypes, and implementation of quality control benchmarks for reproducible differentiation [81]. The ongoing development of maturation strategies to generate more adult-like neurons and glia will better model the aging-related aspects of AD [18]. As these improvements converge, iPSC-based platforms are positioned to become central in the AD drug development pipeline, complementing rather than replacing animal models for specific applications.
Strategic Integration in Research Pipelines
The most effective approach to AD modeling involves strategic integration of multiple model systems based on their complementary strengths. iPSC models excel in human genetic context, patient-specific modeling, and human-specific pathway analysis, while animal models provide essential information on systemic physiology, complex behavior, and whole-organism drug effects. Traditional cell lines remain valuable for initial high-throughput screening and reductionist mechanistic studies. By leveraging the unique advantages of each system at appropriate stages of the research pipeline, the field can accelerate progress toward effective therapies for Alzheimer's disease.
The pursuit of effective Alzheimer's disease (AD) therapeutics faces a critical translational gap, with many promising candidates failing in late-stage clinical trials. Human induced pluripotent stem cell (iPSC) technology has emerged as a transformative platform for disease modeling and preclinical drug screening, offering a human-relevant system that bridges traditional animal models and clinical studies. This whitepaper examines the current state of translational validation for iPSC-based AD models, evaluating their predictive accuracy for clinical trial outcomes. We analyze the technological foundations, present quantitative validation data, detail essential experimental protocols, and identify both capabilities and limitations in the context of drug development pipelines. The integration of iPSC models with advanced technologies like CRISPR gene editing, 3D organoid systems, and machine learning represents a paradigm shift in preclinical validation, potentially accelerating the development of effective AD therapeutics.
Alzheimer's disease drug development has been plagued by a high failure rate, with numerous compounds showing promise in animal models but failing to demonstrate efficacy in human clinical trials. This translational crisis underscores the critical need for more predictive preclinical models that better recapitulate human disease biology [26]. The complex, multifactorial nature of AD—encompassing amyloid-beta accumulation, tau pathology, neuroinflammation, and genetic risk factors—poses particular challenges for traditional model systems [15].
The emergence of iPSC technology over the past decade has revolutionized neurodegenerative disease modeling. By enabling the generation of patient-derived neurons, astrocytes, microglia, and cerebral organoids, iPSC systems provide a human-relevant platform for studying disease mechanisms and screening potential therapeutics [77] [23]. The fundamental premise is that these models capture human-specific biology and genetic diversity in a way that animal models cannot, potentially offering better prediction of clinical outcomes [25]. As of 2025, iPSC-based research has contributed significantly to understanding AD pathophysiology and has begun to influence therapeutic development pipelines, with several candidates identified through iPSC screening advancing toward clinical trials [9].
Foundations of iPSC Technology in Alzheimer's Disease Research
iPSC Generation and Neural Differentiation
The iPSC technology foundation involves reprogramming somatic cells (typically skin fibroblasts or blood cells) into pluripotent stem cells through expression of defined transcription factors, originally OCT4, SOX2, KLF4, and c-MYC [26]. These iPSCs can then be differentially directed toward neural lineages using established protocols that mimic developmental processes:
- Neural Induction: Achieved through dual SMAD signaling inhibition using small molecules like Noggin and SB431542, promoting neuroectodermal specification [23].
- Neural Rosette Formation: Radially organized structures resembling neural tubes appear, serving as reservoirs of neural stem cells (NSCs) [23].
- Cell Type-Specific Differentiation: NSCs can be directed toward specific neuronal subtypes (glutamatergic, GABAergic), astrocytes, or microglia using defined cytokine and growth factor combinations [23] [25].
Key Cellular Models for Alzheimer's Research
Table 1: iPSC-Derived Cellular Models for Alzheimer's Disease Research
| Model Type | Key Features | Applications in AD Research | Limitations |
|---|---|---|---|
| 2D Neuronal Cultures | Cortical neurons expressing relevant AD proteins (APP, tau); amenable to high-content screening | Amyloid and tau pathology studies; synaptic function; high-throughput compound screening | Lack cellular complexity and tissue architecture of human brain |
| iPSC-Derived Astrocytes | Express APOE, clusters; involved in amyloid clearance and inflammatory responses | Studying APOE isoform-specific effects (ε4 vs ε3); neuroinflammation; metabolic support | Limited maturity in standard culture conditions |
| iPSC-Derived Microglia | Express TREM2, CD33; phagocytic activity; inflammatory cytokine production | Neuroinflammation modeling; phagocytosis of amyloid aggregates; microglial-neuronal interactions | Require co-culture with neurons/astrocytes for full activation |
| 3D Cerebral Organoids | Multiple brain region identities; primitive cortical layering; cell-cell interactions more representative of in vivo environment | Modeling amyloid plaque and neurofibrillary tangle formation; network-level dysfunction; neuro-glia interactions | Variable reproducibility; necrotic cores in absence of vascularization |
Quantitative Validation: Comparing iPSC Predictions with Clinical Outcomes
The true test of any preclinical model lies in its ability to predict human clinical outcomes. While the field is still accumulating direct comparative data, several approaches and cases provide insights into the predictive validity of iPSC models.
Drug Repurposing Candidates Identified Through iPSC Screening
Table 2: Drug Repurposing Candidates for AD Identified via iPSC Models
| Compound | Original Indication | iPSC-Based Evidence | Clinical Trial Status/Outcomes | Predictive Accuracy |
|---|---|---|---|---|
| Levetiracetam | Epilepsy | Reduced abnormal neural network activity in AD iPSC-derived neurons | Phase 2: Did not significantly slow cognitive decline in overall population but showed reduced brain atrophy in APOE ε4 non-carriers [1] | Partial: Correctly predicted biological effect but identified subgroup-specific response |
| CT1812 | Novel compound (sigma-2 receptor modulator) | Displaced toxic Aβ and alpha-synuclein oligomers from synapses in iPSC-derived neurons [1] | Phase 2A completed (safety); Phase 2B ongoing for early AD and dementia with Lewy bodies [1] | Pending: Preliminary clinical data appears consistent with proposed mechanism |
| Bosutinib | Chronic myeloid leukemia | Enhanced autophagy and reduced phosphorylated tau levels in iPSC-derived neurons | In clinical trials for amyotrophic lateral sclerosis; AD trials pending [9] | Pending |
Analysis of the current landscape reveals that iPSC models are increasingly being used for lead optimization and safety pharmacology, with at least 25 new drug candidates developed through NIH translational research programs advancing to human trials as of 2025 [1]. These candidates target diverse AD-related biological processes including inflammation, synaptic plasticity, APOE-related mechanisms, and amyloid/tau biology.
Genetic Validation Using CRISPR/Cas9 and iPSC Models
The combination of CRISPR/Cas9 gene editing with iPSC technology provides a powerful approach for validating therapeutic targets and understanding disease mechanisms:
Diagram 1: CRISPR/iPSC Target Validation Workflow
This integrated approach has been successfully applied to several AD-related genes:
- APP/PSEN1/PSEN2: CRISPR correction of familial AD mutations in iPSCs normalizes Aβ42/Aβ40 ratios and reduces phospho-tau levels, validating the amyloid pathway as a therapeutic target [15].
- APOE4: Isoform-specific effects (ApoE4 vs. ApoE3) on Aβ aggregation, lipid metabolism, and neuroinflammation have been confirmed in iPSC-derived astrocytes and microglia, supporting APOE-directed therapeutic approaches [15] [26].
- TREM2: iPSC models with loss-of-function TREM2 mutations recapitulate impaired microglial phagocytosis and metabolic deficits, providing functional validation of this key AD risk gene [25].
Methodological Framework: Experimental Protocols for Translational Validation
Generation of Patient-Specific and Genetically Engineered iPSC Lines
Protocol: Establishing an iPSC Cohort for AD Drug Screening
Donor Selection: Select donors representing AD genetic diversity:
- Familial AD (fAD) with APP, PSEN1, or PSEN2 mutations
- Sporadic AD (sAD) with high and low polygenic risk scores (PRS) [13]
- APOE ε3/ε3, ε3/ε4, and ε4/ε4 genotypes
- Age-matched cognitively healthy controls
Reprogramming:
- Source somatic cells (typically dermal fibroblasts or peripheral blood mononuclear cells)
- Reprogram using non-integrating methods (episomal vectors or Sendai virus) expressing OCT4, SOX2, KLF4, c-MYC, LIN28, and p53 shRNA [23] [26]
- Culture in hypoxic conditions (5% O₂) with ascorbic acid to enhance reprogramming efficiency
CRISPR/Cas9 Genetic Engineering:
- Design sgRNAs targeting AD-relevant genes (APP, APOE, TREM2, etc.)
- Transfert iPSCs with Cas9-sgRNA ribonucleoprotein complexes
- For point mutations (e.g., APOE ε4 to ε3), co-transfect with single-stranded DNA donor templates
- Isolate single-cell clones and validate edits by Sanger sequencing and functional assays [15]
Differentiation into Disease-Relevant Cell Types
Protocol: Directed Differentiation of iPSCs to Cortical Neurons
Neural Induction:
- Culture iPSCs in mTeSR1 medium until 70-80% confluent
- Switch to neural induction medium containing DMEM/F12, N2 supplement, MEM-NEAA, and dual SMAD inhibitors (10 μM SB431542 and 250 ng/mL Noggin) [23]
- Culture for 11-14 days with daily medium changes until neural rosettes appear
Neural Stem Cell (NSC) Expansion:
- Mechanically isolate neural rosettes
- Dissociate and plate on poly-ornithine/laminin-coated surfaces in NSC medium (DMEM/F12, N2, B27, 20 ng/mL FGF2, 20 ng/mL EGF)
- Passage every 5-7 days to maintain NSC cultures
Cortical Neuron Differentiation:
- Plate NSCs at high density (50,000-100,000 cells/cm²) on poly-ornithine/laminin
- Switch to neuronal differentiation medium (Neurobasal, B27, BDNF, GDNF, cAMP, ascorbic acid)
- Culture for 4-8 weeks, with half-medium changes every 3-4 days
- Validate by immunostaining for cortical markers (CTIP2, TBR1, MAP2) and electrophysiological activity [23] [25]
Phenotypic Screening Assays for Therapeutic Evaluation
Protocol: Multi-Parameter Assessment of Compound Efficacy
Amyloid Pathology Assays:
- Aβ ELISAs: Measure Aβ40, Aβ42, and sAPPα/β in conditioned media
- Immunocytochemistry: Quantify amyloid oligomers using conformation-specific antibodies (A11)
Tau Pathology Assays:
- Western Blotting: Assess tau phosphorylation at AD-relevant epitopes (AT8, AT100, PHF1)
- FRET-Based Biosensors: Monitor tau aggregation in live cells
Neuronal Function Assays:
- Calcium Imaging: Measure synchronous network activity using Fluo-4 AM
- Multi-Electrode Arrays: Record spontaneous firing and network bursting
- Synaptic Density: Quantify PSD95 and synapsin puncta by high-content imaging
Neuroinflammation Assays:
- Cytokine Profiling: Measure IL-6, IL-1β, TNF-α in conditioned media
- Microglial Phagocytosis: Assess uptake of pHrodo-labeled Aβ using flow cytometry
Table 3: Key Research Reagents for iPSC-Based Alzheimer's Disease Modeling
| Reagent Category | Specific Examples | Function/Application | Technical Notes |
|---|---|---|---|
| Reprogramming Factors | OCT4, SOX2, KLF4, c-MYC (OSKM); Sendai virus vectors; episomal plasmids | Somatic cell reprogramming to pluripotency | Non-integrating methods preferred for clinical translation |
| Neural Induction Supplements | Noggin; SB431542; LDN-193189; SMAD inhibitors | Directing differentiation toward neural lineage | Dual SMAD inhibition yields >80% PAX6+ neural cells in 11 days [23] |
| Differentiation Factors | FGF2; EGF; BDNF; GDNF; NT-3; NGF | Specific neural subtype specification | Concentrations and timing critical for regional identity |
| CRISPR/Cas9 Components | Cas9 protein; synthetic sgRNAs; HDR templates; nucleofection reagents | Genetic engineering for isogenic controls | RNP complex delivery reduces off-target effects [15] |
| Disease-Relevant Cell Markers | PAX6 (neural progenitors); MAP2 (neurons); GFAP (astrocytes); IBA1 (microglia) | Characterization of differentiated cells | Flow cytometry and immunocytochemistry standard |
| AD Pathology Antibodies | 6E10, 4G8 (Aβ); AT8, PHF1 (p-tau); A11 (oligomers) | Quantifying disease phenotypes | Conformation-specific antibodies preferred for oligomers |
| Functional Assay Reagents | Fluo-4 AM (calcium imaging); FM dyes (synaptic vesicle release); MUSE kits (viability) | Assessing neuronal health and function | High-content imaging enables multiplexed readouts |
Emerging Technologies Enhancing Predictive Validity
Advanced 3D Model Systems
Three-dimensional cerebral organoids and assembloids represent a significant advancement over 2D cultures:
Diagram 2: 3D Organoid Model Development
These systems better recapitulate the cellular complexity and pathological features of AD, including:
- Spontaneous Aβ Plaque Formation: 3D organoids with fAD mutations develop extracellular Aβ deposits after extended culture (2-6 months) [26].
- Neurofibrillary Tangle Pathology: Hyperphosphorylated tau accumulation and tangle-like structures observed in mature organoids [25].
- Neuroinflammation: Microglia-incorporated organoids exhibit disease-associated microglial (DAM) phenotypes and neuroinflammatory responses [25].
Integration with Machine Learning and Computational Approaches
Machine learning algorithms applied to high-content screening data from iPSC models are enhancing predictive accuracy:
- Feature Selection: Genetic algorithms identify optimal predictor combinations from high-dimensional datasets [84].
- Predictive Modeling: LightGBM and other ensemble methods achieve high accuracy (ROC-AUC >0.90) in classifying AD status using multimodal iPSC data [84].
- Pathway Analysis: SHAP (SHapley Additive exPlanations) interpretation identifies non-linear relationships between genetic risk variants and cellular phenotypes [85].
Limitations and Future Directions
Despite their promise, iPSC models face several challenges that impact their predictive validity:
- Developmental Immaturity: iPSC-derived neurons exhibit fetal-like characteristics and may not fully recapitulate aging-related processes central to AD [15] [26].
- Incomplete Pathology: While iPSC models reproduce key molecular features, they rarely develop full amyloid plaques or neurofibrillary tangles without genetic manipulation [25].
- Genetic Complexity: Most iPSC studies focus on monogenic or highly penetrant risk factors, while sAD involves complex polygenic risk architectures [13].
Future directions to enhance translational predictive validity include:
- Incorporation of Polygenic Risk: The IPMAR Resource and similar initiatives are establishing iPSC cohorts representing extremes of AD polygenic risk scores [13].
- Advanced Co-culture Systems: Multi-cell type assembloids incorporating neurons, astrocytes, microglia, and vasculature for studying non-cell-autonomous mechanisms [25].
- Long-term Culture and Progeria Approaches: Extended culture periods and induction of aging-related stressors to accelerate age-associated phenotypes.
- Standardization and Reproducibility: Community standards for differentiation protocols, quality control metrics, and phenotypic assays to enable cross-study comparisons.
iPSC-based models represent a transformative advancement in Alzheimer's disease research, offering unprecedented access to patient-specific human neural cells for therapeutic screening and validation. While the field is still accumulating direct evidence of their predictive validity for clinical trial outcomes, the integration of iPSC technology with CRISPR gene editing, 3D culture systems, and machine learning approaches is steadily closing the translational gap. Current data suggests that iPSC models show particular strength in predicting target engagement and mechanism-based efficacy, though their ability to predict overall clinical outcomes remains partially limited by developmental immaturity and incomplete pathology modeling. As these technologies continue to evolve and standardization improves, iPSC-based screening platforms are poised to become increasingly central to AD drug development pipelines, potentially reducing late-stage attrition rates and accelerating the delivery of effective therapeutics.
The persistent failure of therapeutics developed in traditional animal models to benefit Alzheimer's disease (AD) patients has underscored a critical translational gap in the field, driving the urgent need for more human-relevant disease models [18]. Countless promising therapeutics showing efficacy in rodent AD models have failed in human trials, suggesting that these models do not fully recapitulate the complex pathophysiology of the human condition [18]. This translational challenge has catalyzed the development of two powerful human-based approaches: induced pluripotent stem cell (iPSC) technology, which provides living human neural cells for mechanistic studies and therapeutic screening, and advanced post-mortem brain analysis, which offers a definitive snapshot of end-stage disease pathology in human tissue [18] [86].
The central premise of this technical guide is that the convergence of these two approaches—cross-referencing dynamic molecular signatures from iPSC models with static but authentic pathological data from post-mortem human brains—provides a robust framework for validating disease mechanisms and identifying therapeutic targets with greater translational potential. iPSC technologies have revolutionized preclinical research by enabling the generation of all major brain cell types from patients, including neurons, astrocytes, microglia, and oligodendrocytes, while increasingly complex co-culture systems are being developed to model the intricate cell-cell interactions that occur in the living brain [18] [23]. Simultaneously, advances in post-mortem analysis, including single-cell transcriptomics, proteomics, and multimodal imaging, are providing unprecedented resolution of the molecular and cellular changes in the AD brain [87] [88] [89].
This guide provides researchers with a comprehensive technical framework for conducting rigorous cross-referencing studies between iPSC models and post-mortem brain data, with the goal of establishing pathophysiological concordance—the alignment of disease-associated molecular and cellular signatures between these complementary systems. By systematically validating iPSC-derived findings against the gold standard of human neuropathology, we can enhance the biological relevance of mechanistic studies and accelerate the development of effective therapeutics for this devastating disorder.
Alzheimer's Disease Pathophysiology: Key Pathways and Cellular Players
Core Pathological Hallmarks and Genetic Evidence
Alzheimer's disease is characterized neuropathologically by two defining proteinopathies: extracellular amyloid-β (Aβ) plaques and intracellular neurofibrillary tangles (NFTs) composed of hyperphosphorylated tau protein [18] [90]. These pathological hallmarks are accompanied by progressive neurodegeneration, synaptic dysfunction, gliosis, inflammation, blood-brain barrier (BBB) disruptions, and metabolic disturbances [18]. The amyloid-β pathway remains central to AD pathophysiology, with breakthroughs in molecular medicine positioning it as a crucial therapeutic target [90].
Genetic evidence strongly implicates the Aβ pathway in AD pathogenesis. Early-onset familial AD (FAD), accounting for 1-5% of cases, is caused by highly penetrant autosomal dominant mutations in three genes: the amyloid precursor protein (APP) and presenilin 1 and 2 (PSEN1 and PSEN2), which encode components of the γ-secretase complex that processes APP to generate Aβ peptides [90] [86]. These mutations universally increase the production or aggregation propensity of Aβ42, the longer, more fibrillogenic form of Aβ [86]. For the more common sporadic late-onset AD (SAD), genome-wide association studies (GWAS) have identified numerous risk genes, with APOE4 representing the strongest genetic risk factor, increasing risk approximately 3-fold in heterozygotes and 15-fold in homozygotes [18]. Notably, many more recently identified SAD risk genes are expressed primarily in non-neuronal cells of the brain, shifting research focus toward better understanding the roles of glial cells in neurodegeneration [18].
Beyond Neurons: The Multi-Cellular Nature of AD Pathogenesis
Following years of focus on neuron-based mechanisms, recent genetic and pathological studies have revealed AD as a multi-cellular disorder involving complex interactions between diverse cell types [18] [88]. Oligodendrocytes insulate neuronal axons, astrocytes and microglia form the "tripartite synapse" with neurons and play critical roles in neurotransmitter recycling, metabolic support, and immune surveillance, while the specialized cells of the blood-brain barrier regulate brain homeostasis [18]. Single-cell transcriptomic studies of post-mortem human brains have identified vulnerable populations of both excitatory and inhibitory neurons that are depleted in specific brain regions in AD, as well as glial response states associated with disease progression and cognitive resilience [88].
Recent research has also revealed significant biological heterogeneity within AD, with proteomic analyses of cerebrospinal fluid identifying distinct pathophysiological subtypes characterized by: (1) hyperplasticity and increased BACE1 levels; (2) innate immune activation; and (3) blood-brain barrier dysfunction with low BACE1 levels [91]. These subtypes demonstrate different rates of clinical progression and may require tailored therapeutic approaches, highlighting the importance of capturing this heterogeneity in disease models [91].
Methodological Framework: iPSC Modeling and Post-Mortem Analysis
iPSC Differentiation Strategies for Neural Cell Types
Current protocols for differentiating neural cell types from human iPSCs employ strategies that mimic in vivo developmental steps, progressing through formation of neuroepithelium, specialization of neural stem cells, and differentiation to neural progenitors and mature cell types [23]. These approaches use specific culture media, growth factors, and small molecule inhibitors sequentially added to culture media, with some protocols utilizing inducible overexpression of specific transcription factors to direct differentiation toward particular lineages [23].
Table 1: Key iPSC Differentiation Strategies for Neural Cell Types
| Cell Type | Key Induction Factors | Characteristic Markers | Protocol Duration | Key Applications in AD Research |
|---|---|---|---|---|
| Neural Stem Cells (NSCs) | Dual SMAD inhibition (Noggin, SB431542), FGF2 | PAX6, SOX1, SOX2 | 11-30 days | Intermediate expandable population; foundation for further differentiation |
| Neurons | BDNF, NT-3, GDNF, cAMP, ascorbic acid | MAP2, β-III-tubulin, NeuN | 30-90 days | Study of Aβ toxicity, tau hyperphosphorylation, synaptic dysfunction, neuronal vulnerability |
| Astrocytes | CNTF, BMPs, LIF, FBS | GFAP, S100β, AQP4 | 60-120 days | Role in tripartite synapse, metabolic support, inflammatory response, Aβ clearance |
| Microglia | IL-34, CSF-1, GM-CSF, TGF-β | TMEM119, P2RY12, IBA1 | 30-60 days | Innate immune activation, phagocytosis of Aβ, neuroinflammation, DAM phenotype |
| Cerebral Organoids | Matrigel, neural induction followed by spontaneous differentiation | Mixed regional and cell type markers | 30-180+ days | Complex cell-cell interactions, tissue-level organization, disease progression in 3D context |
The initiation of neural differentiation in iPSCs typically leads to formation of neural rosettes—radially organized cellular structures regarded as an early stage of neural development in vitro that function as a reservoir of neural stem cells (NSCs) [23]. A highly reliable protocol to generate NSCs introduces dual inhibition of SMAD signaling in iPSCs as the initial step, using Noggin and SB431542 together with specific initial cell density to generate a high number of PAX6+ neural cells competent of rosette formation within 11 days [23]. These NSCs can be isolated and propagated in vitro while maintaining their self-renewing capacity and differentiation potential to both neuronal and glial cell types [23].
For specific neuronal and glial differentiation, numerous protocols have been optimized. Neuronal differentiation typically employs factors such as BDNF, NT-3, GDNF, cAMP, and ascorbic acid over 30-90 days to generate mature, functional neurons [23] [86]. Astrocyte differentiation requires longer timelines (60-120 days) using CNTF, BMPs, LIF, or serum-containing media, while microglia differentiation utilizes IL-34, CSF-1, GM-CSF, and TGF-β over 30-60 days [23] [25]. Additionally, 3D cerebral organoid models provide more functionally complex systems that better mimic developmental processes and enable study of complex cell-cell interactions in a tissue-like context [23].
Advanced Post-Mortem Brain Analysis Techniques
Modern post-mortem brain analysis has evolved far beyond traditional histology to incorporate multi-omics approaches at single-cell resolution, providing unprecedented insights into the molecular landscape of the AD brain. Recent advances include:
Single-cell and single-nucleus RNA sequencing enables comprehensive characterization of cellular diversity and transcriptomic alterations across brain regions and individuals. A landmark 2024 study profiled over 1.3 million nuclei from six brain regions (entorhinal cortex, hippocampus, anterior thalamus, angular gyrus, midtemporal cortex, and prefrontal cortex) from 48 individuals, identifying 76 cell types, including region-specific subtypes of astrocytes and excitatory neurons, and revealing vulnerable neuronal populations depleted in AD [88].
Proteomic analyses provide crucial insights not always detectable at the RNA level. A large-scale deep multi-layer analysis of Alzheimer's disease brain revealed strong proteomic disease-related changes not observed at the RNA level, identifying 44 groups of proteins ("protein communities") that showed coordinated changes in AD brains [89]. Two key communities not detected at the RNA level were associated with MAPK signaling/energy metabolism and early disease stages when amyloid and tau begin to accumulate [89].
Multimodal imaging-pathology correlation approaches, such as those used in the South Texas Alzheimer's Disease Research Center repository, employ advanced MRI sequences followed by MRI-guided histopathological examination to draw exact correspondences between neuroimaging markers and pathological findings [87]. This repository includes 139 post-mortem brains with a wide range of neurodegenerative disorders, specifically designed to disentangle the effects of comorbid conditions, with only 3% showing evidence of AD-only pathology while 76% had multiple pathologies contributing to clinical diagnosis [87].
Table 2: Key Analytical Approaches for Post-Mortem Brain Tissue
| Analytical Method | Key Outputs | Technical Considerations | Applications in AD Research |
|---|---|---|---|
| Single-cell/single-nucleus RNA-seq | Cell type identification, differential expression, cellular abundance changes | Requires high-quality tissue with minimal RNA degradation; computational expertise for data analysis | Identification of vulnerable and resilient cell populations; regional differences in transcriptomic responses [88] |
| Proteomics (mass spectrometry) | Protein abundance, post-translational modifications, protein-protein interactions | Can reveal changes not detectable at RNA level; requires specialized instrumentation | Discovery of disease-associated protein communities and pathways [89] |
| Multiplexed immunofluorescence | Spatial localization of multiple cell types and markers in tissue context | Limited by antibody quality and availability; enables spatial analysis | Mapping of pathological protein spread; cell-cell interactions in situ |
| MRI-guided histopathology | Correlation of in vivo imaging markers with pathological ground truth | Requires specialized equipment and protocols for tissue processing | Validation of neuroimaging biomarkers; lesion-specific analysis [87] |
| CSF proteomics | Identification of pathophysiological subtypes, biomarker discovery | Reflects global brain changes; accessible in living patients | Subtype stratification (hyperplasticity, innate immune activation, BBB dysfunction) [91] |
Establishing Concordance: Cross-Referencing Methodologies
Experimental Design for Cross-Validation Studies
Rigorous experimental design is essential for meaningful cross-referencing between iPSC models and post-mortem data. Several key considerations must be addressed:
Cohort Selection and Matching: When comparing to post-mortem datasets, ensure appropriate matching of relevant genetic backgrounds (APOE genotype, specific risk variants), clinical phenotypes, and pathological severity. For studies targeting specific AD subtypes identified through proteomic profiling (hyperplasticity, innate immune activation, or BBB dysfunction), select iPSC lines that correspond to these subtypes when possible [91].
Regional Specificity: Account for the well-established regional vulnerability in AD pathology progression. When modeling specific stages of disease, select brain regions corresponding to appropriate Braak stages: entorhinal cortex for early stages (I-II), hippocampus and thalamus for mid-stages (III-IV), and neocortical regions for late stages (V-VI) [88]. Ensure iPSC-derived cells represent appropriate regional identities when comparing to region-specific post-mortem data.
Multi-modal Data Integration: Develop analysis plans that incorporate multiple data types (transcriptomic, proteomic, epigenetic, functional) from both systems to establish concordance across different molecular layers. Prioritize direct measurement of proteins alongside RNA when possible, as proteomic changes may reveal important disease-related alterations not detectable at the RNA level [89].
Longitudinal Design: For iPSC studies, incorporate temporal dimensions that capture disease progression rather than single endpoints, as AD unfolds over decades in humans. This enables better alignment with the cross-sectional but stage-rich post-mortem data.
Analytical Approaches for Data Integration
Several computational approaches facilitate systematic comparison between iPSC and post-mortem datasets:
Differential Expression Concordance Analysis: Identify genes/proteins differentially expressed in both systems and assess the direction and magnitude of correlation. More sophisticated approaches can evaluate whether entire gene modules or pathways show consistent alterations.
Cell Type Deconvolution: Use signature gene sets from purified post-mortem cell types to assess the similarity and maturity of iPSC-derived cells. Single-cell reference atlases from post-mortem brains [88] provide excellent resources for benchmarking iPSC-derived cellular models.
Network Biology Approaches: Construct co-expression networks or protein-protein interaction networks in both systems and identify preserved modules associated with disease. The "protein communities" concept from proteomic studies of post-mortem tissue [89] can be applied to iPSC proteomic data to identify conserved functional units.
Pathway Enrichment Analysis: Use gene set enrichment analysis (GSEA) and related methods to identify biological pathways consistently altered in both systems, focusing on pathways with established relevance to AD such as those related to synaptic function, immune response, protein homeostasis, and metabolism.
Key Findings from Concordance Studies
Validated Cellular and Molecular Signatures
Cross-referencing studies have yielded several important validated findings that demonstrate the power of this convergent approach:
Vulnerable Neuronal Populations: Single-nucleus RNA sequencing of post-mortem brains has identified specific excitatory neuron subtypes in the entorhinal cortex that are preferentially vulnerable in early AD [88]. iPSC-derived neurons carrying AD risk variants have confirmed intrinsic vulnerabilities in these neuronal subtypes and implicated the Reelin signaling pathway in modulating neuronal vulnerability [88].
Disease-Associated Microglia (DAM): Post-mortem analyses have identified a characteristic microglial state associated with AD, known as disease-associated microglia (DAM) [25]. iPSC-derived microglia with AD risk variants in genes such as TREM2 have recapitulated key aspects of this phenotype, including altered phagocytic capacity and lipid metabolism, validating the relevance of iPSC models for studying neuroimmune mechanisms in AD [25].
Astrocyte Heterogeneity and Resilience Signatures: Recent multiregion single-cell atlas of AD identified an astrocyte transcriptional program associated with cognitive resilience to AD pathology, linking choline metabolism and polyamine biosynthesis in astrocytes to preserved cognitive function late in life [88]. iPSC-derived astrocytes from cognitively resilient individuals provide a model system for mechanistically dissecting these protective pathways.
Proteomic Alterations in MAPK and Metabolic Pathways: Large-scale proteomic analysis of post-mortem brains revealed disease-associated changes in protein communities related to MAPK signaling and energy metabolism that were not detected at the RNA level [89]. Follow-up studies in iPSC models can now validate the functional significance of these pathway alterations and test therapeutic strategies targeting these processes.
Regional Vulnerability Patterns
The systematic regional analysis of post-mortem tissue has revealed striking patterns of cellular vulnerability that can be modeled in region-specific iPSC-derived cultures:
Entorhinal Cortex Vulnerability: Excitatory neurons in superficial layers of the entorhinal cortex show early vulnerability to tau pathology and neurodegeneration [88]. iPSC-derived neurons with entorhinal identity can model the cell-autonomous and non-cell-autonomous mechanisms underlying this regional vulnerability.
Thalamic Inhibitory Neuron Specialization: The thalamus contains a unique inhibitory neuron population (MEIS2+FOXP2+) distinct from canonical cortical inhibitory subtypes [88]. This regional specialization may contribute to differential vulnerability and represents an important consideration when modeling thalamic involvement in AD.
Hippocampal Subfield Vulnerabilities: Excitatory neuron subtypes across hippocampal subfields (CA1, CA2/CA3, dentate gyrus) show differential vulnerability that can be modeled in iPSC-derived cultures with specific hippocampal identities [88].
Table 3: Key Research Reagent Solutions for Concordance Studies
| Reagent/Resource Category | Specific Examples | Application in Concordance Studies | Technical Notes |
|---|---|---|---|
| iPSC Lines | FAD-mutant lines (APP, PSEN1, PSEN2), SAD patient-derived lines, isogenic control lines | Modeling genetic risk factors; establishing causality through genetic correction | Prioritize lines with comprehensive genetic and clinical annotation; use isogenic controls for confident attribution of phenotypes |
| Differentiation Kits & Reagents | SMAD inhibitors (Noggin, SB431542), patterning factors (FGF2, BDNF, GDNF), cell type-specific media | Generation of specific neural cell types with regional identities | Maintain rigorous quality control; validate differentiation efficiency with multiple markers; use same differentiation protocols for cross-study comparisons |
| Antibodies for Validation | Cell identity markers (PAX6, MAP2, GFAP, TMEM119), pathological markers (Aβ, p-tau), functional markers (synaptophysin, PSD95) | Characterizing iPSC-derived cells; comparing protein expression to post-mortem tissue | Validate antibodies in both iPSC-derived cells and post-mortem tissue; use multiplexed approaches for limited samples |
| Post-Mortem Reference Datasets | Single-cell atlases [88], proteomic datasets [89], imaging-pathology repositories [87] | Benchmarking iPSC models; identifying conserved signatures | Use multiple independent datasets for validation; account for technical differences in sample processing |
| Bioinformatics Tools | Cell type deconvolution algorithms (CIBERSORTx), single-cell analysis tools (Seurat, Scanpy), pathway analysis tools (GSEA) | Computational integration of iPSC and post-mortem data | Standardize analytical pipelines; use appropriate statistical corrections for multiple comparisons |
| 3D Culture Systems | Cerebral organoids, assembloids, microfluidic devices | Modeling complex cell-cell interactions and tissue-level pathology | Improved physiological relevance but increased technical complexity and variability |
Visualizing Workflows and Pathways
Experimental Workflow for Concordance Studies
Key Signaling Pathways in AD Pathophysiology
The systematic cross-referencing of iPSC molecular signatures with post-mortem human brain data represents a powerful approach for advancing our understanding of Alzheimer's disease mechanisms and developing effective therapeutics. As both technologies continue to evolve, several exciting directions emerge:
Advanced 3D Model Systems: The development of more complex cerebral organoids and assembloids that better recapitulate the cellular diversity and spatial organization of the human brain will enable more accurate modeling of region-specific vulnerabilities and cell-cell interactions observed in post-mortem tissue [23].
Spatial Multi-omics Integration: Emerging spatial transcriptomics and proteomics technologies applied to both iPSC models and post-mortem tissue will enable direct comparison of spatial patterns of gene expression and pathology progression, bridging a critical gap between conventional 2D cultures and intact brain tissue.
Longitudinal Modeling of Disease Progression: Incorporating temporal dimensions into iPSC models through inducible gene expression systems, chronic treatment paradigms, and repeated sampling will better capture the progressive nature of AD and enable alignment with cross-sectional post-mortem data representing different disease stages.
Personalized Medicine Approaches: Leveraging iPSC banks from individuals with extensive clinical, imaging, and ultimately neuropathological data will enable truly personalized modeling and help decipher the biological basis of heterogeneity in AD presentation and progression [91].
In conclusion, the pathophysiological concordance framework provides a rigorous approach for validating disease mechanisms and therapeutic targets in human-relevant systems. By systematically cross-referencing dynamic molecular signatures from iPSC models with the gold standard of post-mortem neuropathology, researchers can enhance the translational predictive value of preclinical studies and accelerate the development of effective therapies for this devastating disorder. As these technologies continue to advance and converge, they offer renewed hope in tackling the complex challenge of Alzheimer's disease.
The Emergence of Clinical Trials Informed by iPSC Research (e.g., Bosutinib, Ropinirole, Bromocriptine)
The high failure rate of clinical trials for neurodegenerative diseases underscores an urgent need for improved disease models that can better predict clinical efficacy in humans [67]. The advent of induced pluripotent stem cell (iPSC) technology has revolutionized preclinical drug discovery by providing access to patient-derived neural cells that recapitulate the complex genomic background of human diseases [9] [92]. This whitepaper examines the emerging paradigm of iPSC-informed clinical trials, focusing on several drug candidates that have transitioned from iPSC-based screening platforms to human trials for Alzheimer's disease (AD) and other neurodegenerative conditions. By leveraging patient-specific cellular models, this approach enables more physiologically relevant screening of compound libraries and better assessment of therapeutic efficacy and toxicity before advancing to clinical trials [93] [92].
The integration of iPSC technology into the drug development pipeline represents a significant advancement for addressing Alzheimer's disease, a condition characterized by progressively declining cognitive function and irreversible neurodegeneration [67]. With over 46 million people affected worldwide and no disease-modifying treatments currently available, the need for innovative approaches is particularly pressing [67]. This technical guide explores how iPSC-based models are being utilized to elucidate AD mechanisms and identify repurposed drug candidates with novel mechanisms of action against neurodegenerative pathology.
iPSC Technology in Neurodegenerative Disease Research
Fundamentals of iPSC-Derived Neural Models
Induced pluripotent stem cells are generated by reprogramming somatic cells (typically skin fibroblasts or blood mononuclear cells) to a pluripotent state through the forced expression of specific transcription factors, originally Oct4, Sox2, cMyc, and Klf4 [67] [94]. These reprogrammed cells can then be differentiated into various neural cell types, including cortical neurons, astrocytes, microglia, and oligodendrocytes, or more complex 3D organoid systems that better mimic the brain's cellular architecture [67] [93] [95].
The differentiation process typically follows developmental principles, beginning with the formation of neuroepithelium and neural stem cells (NSCs), which subsequently specialize into specific neural lineages [93]. Protocols often employ dual SMAD signaling inhibition using compounds like Noggin and SB431542 to efficiently direct differentiation toward neural fates [93]. These approaches can generate relatively homogeneous populations of neural cells that retain the genetic signatures of the original patient, making them particularly valuable for modeling both familial and sporadic forms of neurodegenerative diseases [94] [93].
Advantages of iPSC Models for Alzheimer's Disease Research
iPSC-based models offer several critical advantages for Alzheimer's disease research:
Patient-specific pathophysiology: iPSCs can be generated from both familial AD (FAD) patients with mutations in APP, PSEN1, or PSEN2 genes, and sporadic AD (SAD) patients with complex genetic risk factors [67] [94]. This enables researchers to study disease mechanisms across different AD subtypes.
Human-relevant systems: Unlike rodent models that may not fully recapitulate human disease due to species-specific differences in protein expression and regulation, iPSC-derived neural cells provide a human genomic context for studying disease pathways and therapeutic responses [67] [95].
Functional assessment capability: iPSC-derived neurons can be cultured on microelectrode arrays (MEAs) to measure electrophysiological properties and long-term potentiation (LTP), providing functional readouts of synaptic health and network activity in response to pathological insults and therapeutic interventions [96].
Multi-cell type interactions: Developing co-culture systems allow researchers to model the complex interactions between neurons, astrocytes, microglia, and other brain cell types that contribute to AD progression [95].
iPSC-Informed Drug Candidates in Clinical Development
The application of iPSC-based screening platforms has identified several repurposed drug candidates that have advanced to clinical trials for neurodegenerative diseases. The table below summarizes key candidates with their mechanisms of action and current development status.
Table 1: Drug Candidates Identified Through iPSC Screening Advancing to Clinical Trials
| Drug Candidate | Original Indication | Proposed Mechanism in Neurodegeneration | Current Trial Status | Key Supporting iPSC Findings |
|---|---|---|---|---|
| Bosutinib | Chronic myeloid leukemia | Inhibits Src/c-Abl pathway, enhances autophagy, reduces protein aggregates [97] [92] | Clinical trials for ALS [9] [97] | Reduces phosphorylated TDP-43, improves motor neuron survival, extends survival in ALS models [97] [92] |
| Ropinirole | Parkinson's disease | Dopamine D2 receptor agonist, inhibits oxidative stress [97] [92] | Clinical trials for ALS [9] [33] [97] | Suppresses oxidative stress in patient-derived motor neurons [97] [92] |
| Bromocriptine | Parkinson's disease, hyperprolactinemia | Reduces Aβ production through unknown mechanism [92] | Preclinical/Clinical development for AD [92] | Identified as most effective compound at reducing Aβ in iPSC-derived neuron screening [92] |
| Ezogabine (Retigabine) | Epilepsy | Potassium channel opener, reduces neuronal hyperexcitability [97] | Clinical trials for ALS [9] [33] [97] | Normalizes hyperexcitability of patient-derived cortical and spinal motor neurons [97] |
Bosutinib: Targeting Protein Aggregation Pathways
Bosutinib, an Src/c-Abl inhibitor originally developed for chronic myeloid leukemia, was identified through a screening of 1,416 approved drugs using an iPSC-derived induced motor neuron (iMN) system [92]. The screening platform utilized overexpression of transcription factors Lhx3, Ngn2, and Isl1 to generate functional motor neurons from patient-derived iPSCs [92]. Bosutinib demonstrated significant effects in enhancing autophagic flux, reducing protein aggregates including phosphorylated TDP-43, improving motor neuron survival, and extending survival in ALS mouse models [97] [92].
Subsequent clinical investigation through the iDReAM study showed that ALS patients with lower baseline levels of neurofilament light chain (a biomarker of axonal injury) responded better to bosutinib treatment, with slowed functional decline [92]. Importantly, the Src/c-Abl pathway targeted by bosutinib has also shown promise in Alzheimer's and Parkinson's disease models, suggesting it may represent a common molecular target across multiple neurodegenerative conditions characterized by protein aggregation [92].
Ropinirole and Bromocriptine: Dopaminergic Agonists with Novel Mechanisms
Ropinirole, a dopamine D2 receptor agonist used for Parkinson's disease, was identified as a candidate anti-ALS therapeutic through iPSC-based drug screening [97]. Interestingly, its potential anti-ALS mechanism appears to be independent of its antioxidant activity or dopamine D2 receptor agonism, suggesting a novel pathway yet to be fully elucidated [97]. In iPSC-derived motor neuron models, ropinirole demonstrated significant protective effects against oxidative stress, a key pathophysiological mechanism in ALS [97] [92].
Bromocriptine, another dopaminergic agonist, was identified as the most effective compound for reducing Aβ production in a screening of existing drugs using iPSC-derived cortical neurons [92]. The screening approach quantified Aβ secretion as a primary readout, and bromocriptine emerged as the lead candidate from this phenotypic screen. Based on these iPSC-derived data, bromocriptine is now being advanced toward clinical trials for Alzheimer's disease [92].
Experimental Protocols for iPSC-Based Drug Screening
Neural Differentiation and Disease Modeling
The standard workflow for iPSC-based drug discovery involves several critical steps, each requiring optimized protocols and quality control measures:
iPSC Generation and Maintenance:
- Source somatic cells (typically skin fibroblasts or peripheral blood mononuclear cells) from patients and healthy controls [67] [93]
- Reprogram using non-integrating Sendai virus or episomal vectors expressing OCT4, SOX2, KLF4, and c-MYC [67] [93]
- Culture on feeder layers or in defined, feeder-free conditions with essential supplements like bFGF to maintain pluripotency [93]
- Regularly validate pluripotency markers (OCT4, NANOG, SSEA-4) and karyotype stability [93]
Neural Induction and Differentiation:
- For cortical neurons: Use dual SMAD inhibition with Noggin (or LDN-193189) and SB431542 for efficient neural induction [93] [96]
- Pattern toward cortical fate using sequential treatment with FGF2, WNT antagonists, and BDNF [93] [96]
- For astrocytes: Extended differentiation (60-90 days) with CNTF, BMPs, or LIF to promote glial fate [93]
- For 3D models: Use embryoid body formation or matrix embedding with progressive patterning factors [93]
Disease Phenotype Induction:
- For familial AD models: Utilize CRISPR/Cas9 to introduce specific mutations (APP, PSEN1, PSEN2) into control iPSCs [67] [94]
- For sporadic AD modeling: Use extended maturation (90-180 days) to mimic age-related changes [96] [95]
- Acute phenotype induction with Aβ42 oligomers (typically 1-5 μM for 24-72 hours) to model synaptic toxicity [96]
Table 2: Key Reagent Solutions for iPSC-Based Alzheimer's Disease Research
| Research Reagent | Function | Example Application in AD Models |
|---|---|---|
| Noggin / LDN-193189 | BMP pathway inhibitor, neural induction | Dual SMAD inhibition for efficient neural differentiation [93] |
| SB431542 | TGF-β pathway inhibitor, neural induction | Combined with Noggin for neural specification [93] |
| BDNF, GDNF, NT-3 | Neurotrophic factors | Enhance neuronal survival and maturation [93] [92] |
| Aβ42 oligomers | Induce AD-related pathology | Acute treatment to model synaptic dysfunction and tau phosphorylation [96] |
| Microelectrode arrays (MEAs) | Electrophysiological assessment | Measure neuronal network activity, LTP, and drug effects on synaptic function [96] |
| CRISPR/Cas9 systems | Genome editing | Introduce FAD mutations or correct risk alleles in patient iPSCs [67] [94] |
Compound Screening and Validation
The drug screening process typically involves both targeted and unbiased approaches:
High-Content Phenotypic Screening:
- Plate iPSC-derived neurons in 384-well formats for high-throughput imaging
- Treat with compound libraries (typically 1-10 μM concentration range)
- Assess multiple parameters: neuronal survival, synaptic density (synapsin-1, PSD-95), phospho-tau levels, and apoptotic markers [96] [92]
- For Aβ-focused screens, measure Aβ40/Aβ42 ratios via ELISA or immunostaining [92]
Functional Assessment Using Microelectrode Arrays:
- Plate cortical neurons on MEAs pre-treated with patterning agents (PEG, DETA) to guide network formation [96]
- Record baseline electrical activity and long-term potentiation (LTP) induced by high-frequency stimulation (500 mV, 5 ms) [96]
- Apply test compounds before or after Aβ42 oligomer challenge
- Quantify changes in firing rate, synchrony, and LTP maintenance as indicators of functional rescue [96]
Mechanistic Validation:
Diagram 1: iPSC Drug Screening Workflow for Alzheimer's Disease
Signaling Pathways and Mechanisms of Action
The drug candidates identified through iPSC screening target diverse molecular pathways involved in Alzheimer's disease pathogenesis. Understanding these mechanisms provides insights into both the compounds' therapeutic potential and the underlying biology of neurodegeneration.
Src/c-Abl Pathway in Protein Aggregation
Bosutinib's identification highlighted the importance of the Src/c-Abl signaling pathway in neurodegenerative protein aggregation. In healthy neurons, this pathway regulates various cellular processes including cytoskeletal organization and vesicle trafficking [92]. In AD and ALS models, elevated Src/c-Abl activity contributes to pathological hyperphosphorylation of proteins including tau and TDP-43, promoting their aggregation and impairing clearance mechanisms [92].
iPSC-derived neurons from patients with sporadic AD show increased activation of this pathway, which can be normalized by bosutinib treatment. The drug enhances autophagic clearance of protein aggregates through inhibition of c-Abl, reducing the accumulation of phosphorylated tau and other pathological proteins [92]. This mechanism represents a promising approach across multiple neurodegenerative conditions, as protein aggregation is a common feature of these diseases.
Neuronal Hyperexcitability and Network Dysfunction
Retigabine (ezogabine) targets a fundamentally different aspect of neurodegeneration – neuronal hyperexcitability. iPSC-based studies have revealed that neurons from both ALS and AD patients exhibit abnormal firing patterns and network hyperexcitability that contributes to excitotoxicity and accelerated neurodegeneration [97] [96].
Retigabine acts as a potassium channel opener, specifically activating KCNQ (Kv7) channels to reduce neuronal excitability and restore normal firing patterns [97]. In iPSC-derived cortical neurons, retigabine treatment normalized Aβ42-induced hyperexcitability and improved neuronal survival [97] [96]. This approach demonstrates how iPSC models can identify pathophysiology that may not be apparent in traditional animal models and highlight novel therapeutic targets.
Diagram 2: Therapeutic Targets in Alzheimer's Disease Pathogenesis
The emergence of clinical trials informed by iPSC research represents a paradigm shift in neurodegenerative disease drug development. Candidates like bosutinib, ropinirole, and bromocriptine demonstrate how patient-specific cellular models can identify repurposing opportunities with novel mechanisms of action against Alzheimer's disease pathology. The integration of iPSC-based screening with functional validation using electrophysiological approaches and multi-omics analyses provides a powerful platform for de-risking drug candidates before advancing to clinical trials.
As these technologies continue to evolve—through improved differentiation protocols, more complex 3D model systems, and integration with artificial intelligence—iPSC-informed drug discovery is poised to play an increasingly central role in overcoming the high failure rates that have plagued neurodegenerative disease therapeutics. The ongoing clinical trials for bosutinib, ropinirole, and other iPSC-identified compounds will provide critical validation of this approach and potentially deliver the first disease-modifying treatments informed by human iPSC models.
Leveraging Artificial Intelligence (AI) for Multi-Omics Data Integration and Enhanced Phenotypic Prediction
Alzheimer's disease (AD) remains a profound public health challenge, characterized by progressive cognitive decline and the absence of curative treatments [12]. The multifactorial nature of AD, shaped by complex interactions between genetic, environmental, and biological factors, has complicated both research and clinical management [98]. Current models often fail to fully replicate human physiology and pathology, creating a critical need for more human-relevant research platforms [12].
The convergence of human induced pluripotent stem cell (iPSC) technology, multi-omics technologies, and artificial intelligence (AI) is creating a transformative paradigm in AD research [98] [12]. iPSC models capture patient-specific genetic backgrounds, enabling the generation of neurons, glia, and organoids that retain donor-specific molecular signatures [12]. When combined with AI-driven analytical frameworks capable of integrating large-scale genomic, transcriptomic, proteomic, metabolomic, and microbiomic datasets, these models provide unprecedented opportunities to elucidate molecular mechanisms, identify early biomarkers, and accelerate therapeutic development [98] [99].
This technical guide examines cutting-edge methodologies at the intersection of AI, multi-omics integration, and iPSC-based disease modeling, providing researchers with practical frameworks for enhancing phenotypic prediction in Alzheimer's disease research.
Computational Foundations of AI-Driven Multi-Omics Integration
Machine Learning Approaches for Data Integration
AI-driven multi-omics analysis leverages sophisticated computational approaches to integrate diverse molecular datasets. Machine learning and deep learning algorithms have demonstrated particular utility in managing the complexity, high dimensionality, and heterogeneity of multi-omics data [98]. These include:
- Deep belief networks and joint deep semi-non-negative matrix factorization for disease classification and patient stratification [98]
- Network-based models including protein-protein interaction (PPI) networks reconstructed with centrality analysis to identify key hub genes [99]
- Multimodal foundation models that connect molecular structures with phenotypic information using dual-space contrastive learning frameworks [100]
These approaches enable researchers to move beyond single-omics analyses toward a systems biology perspective that more accurately reflects the complexity of Alzheimer's disease pathophysiology.
Key Analytical Outputs in Alzheimer's Research
AI-driven multi-omics analyses have yielded several critical insights into AD mechanisms and potential biomarkers:
Table 1: Key Biomarkers and Pathways Identified Through AI-Driven Multi-Omics Analysis in AD
| Analysis Type | Identified Biomarkers/Pathways | Biological Significance | Reference |
|---|---|---|---|
| Proteomics & Transcriptomics | APP, YWHAE, YWHAH, SOD1, UQCRFS1, ATP5F1B, AP2M1, MMAB, INA, RPL6, HADHB, CD63, CTNNB1 | Key hub genes implicated in both early and advanced AD | [99] |
| Pathway Analysis | Oxidative phosphorylation, metabolic pathways, synaptic transmission | Critical pathways associated with AD progression | [99] |
| Single-cell Transcriptomics | Disease-associated microglia (DAM), A1/A2 astrocyte subtypes, inflammatory signaling | Cellular heterogeneity in neuroinflammation | [101] |
| Functional Genomics | TREM2, SPP1, CX3CR1, APOE, BINI, PICALM | Microglial function in Aβ clearance and neuroinflammation | [101] |
Experimental Frameworks for iPSC-Based AD Modeling
iPSC Resource Development and Characterization
The establishment of robust iPSC resources is foundational to effective AD modeling. Recent large-scale initiatives have demonstrated several critical methodological considerations:
Donor Selection: Donors should be selected from well-characterized cohorts with extensive clinical, longitudinal, and genetic datasets. The IPMAR Resource exemplifies this approach, selecting donors from a UK cohort of 6,000+ research-diagnosed early or late-onset AD cases and elderly cognitively healthy controls who have lived through the age of risk for disease development (>85 years) [102].
Genetic Risk Stratification: Capture extremes of polygenic risk through global AD polygenic risk scores (PRS) and pathway-specific genetic risk (e.g., complement pathway) [102]. The iDA Project provides 200 iPSC lines representing nearly all common APOE genotypes from the Alzheimer's Disease Neuroimaging Initiative (ADNI) participants [103].
Standardized Characterization: Implement rigorous quality control including pluripotency validation, karyotyping, and microbiological testing. Ensure associated datasets include comprehensive clinical, imaging, and genetic information [102] [103].
Differentiation and Phenotypic Screening Protocols
The transition from iPSCs to relevant neural cell types requires optimized differentiation protocols and phenotypic screening approaches:
Neural Differentiation: Adapt established protocols for generating cortical neurons, microglia, and astrocytes from iPSCs. Incorporate 3D organoid systems to better model tissue-level complexity and cell-cell interactions [12] [104].
High-Content Phenotypic Screening: Implement high-content imaging platforms capable of capturing thousands of cellular images across multiple wells and plates. Key parameters include [105]:
- Adjust exposure time to avoid overexposed images
- Set correct offset from autofocus to ensure proper focus
- Capture optimal number of images per well to adequately represent cell population
Multiplexed Assays: Utilize approaches like Cell Painting that employ multiple fluorescent dyes to stain distinct cellular components, converting cellular images into quantifiable data for computational analysis [105] [104].
AI Methodologies for Enhanced Phenotypic Prediction
Image Analysis and Feature Extraction
AI-powered image analysis represents a critical advancement in extracting meaningful information from high-content phenotypic screens:
Segmentation and Feature Extraction: Utilize tools like CellProfiler or KNIME for segmenting individual cells and extracting hundreds of features covering shape, texture, intensity, and granularity [105]. Advanced platforms like Ardigen's phenAID leverage state-of-the-art computer vision and deep learning to extract high-dimensional features from high-content screening images [105].
Deep Learning Approaches: Implement convolutional neural networks (CNNs) for processing high-content images and recognizing intricate phenotypic patterns [104]. These excel at detecting subtle morphological variations in drug-treated cells that may escape human observation [104].
Data Quality Control: Maintain rigorous quality control standards throughout image analysis. Monitor for blurred images, debris, contamination, and ensure cell count matches expectations [105].
Multimodal Integration for Phenotypic Prediction
Advanced AI models now enable integration of morphological data with multi-omics datasets:
Foundation Models: Develop multimodal foundation models like PhenoModel, which uses dual-space contrastive learning to connect molecular structures with phenotypic information [100]. These models are applicable to various downstream tasks including molecular property prediction and active molecule screening.
Phenotypic Profiling: Leverage platforms that integrate phenotypic profiles with omics data and chemical structures to enhance prediction accuracy and robustness [105]. This integration allows for identification of correlations between drug-induced perturbations and cellular behavior.
Table 2: AI Approaches for Phenotypic Prediction in Alzheimer's Disease Research
| AI Methodology | Application in AD Research | Key Advantages | Implementation Considerations | |
|---|---|---|---|---|
| Deep Learning/Computer Vision | Analysis of high-content imaging from iPSC-derived cells | Detects subtle morphological changes; unbiased analysis | Requires large, annotated datasets; computationally intensive | [105] [104] |
| Multimodal Foundation Models | Connecting molecular structures with phenotypic information | Enables molecular property prediction and virtual screening | Dependent on quality of both chemical and phenotypic data | [100] |
| Network-Based Models | Protein-protein interaction network analysis | Identifies key hub genes and pathways | Requires high-quality interaction databases | [98] [99] |
| Generative Adversarial Networks (GANs) | Novel compound design for desired phenotypic outcomes | Generates novel molecular structures with optimized properties | Outputs require experimental validation | [106] |
The Scientist's Toolkit: Essential Research Reagent Solutions
Table 3: Key Research Reagent Solutions for AI-Driven Multi-Omics and Phenotypic Screening
| Research Tool | Function/Application | Key Features | Representative Examples |
|---|---|---|---|
| iPSC Repository Banks | Provide characterized iPSC lines for AD research | Sex-balanced, multiple APOE genotypes, associated clinical data | iDA Project [103], IPMAR Resource [102] |
| High-Content Imaging Systems | Automated microscopy for phenotypic screening | Multiplexed channels, automated acquisition, high throughput | CellXpress.ai [104] |
| Image Analysis Software | Segmentation and feature extraction from cellular images | Deep learning algorithms, high-dimensional feature extraction | Ardigen phenAID [105], CellProfiler [105] |
| Multi-Omics Analytics Platforms | Integration of transcriptomic, proteomic, and other omics data | AI-driven integration, network analysis, biomarker discovery | PhenoModel [100] |
| 3D Cell Culture Systems | Physiologically relevant models for phenotypic screening | Better mimic tissue environment, organoid development | Organ-on-a-chip technologies [104] |
Future Directions and Implementation Challenges
Addressing Current Limitations
Despite promising advances, several challenges remain in fully realizing the potential of AI-driven multi-omics integration:
Data Heterogeneity: Inconsistencies in data processing methods across platforms and laboratories complicate integration efforts [98]. Implementing FAIR (Findable, Accessible, Interoperable, Reusable) principles from project inception can enhance reproducibility and integration [105].
Model Interpretability: The "black box" nature of some complex AI models limits biological insight and clinical adoption [98] [104]. Developing explainable AI approaches and maintaining domain expert oversight throughout analysis is crucial [105].
Clinical Validation: Many identified biomarkers and pathways require experimental validation in physiologically relevant systems [98] [99]. Advanced iPSC models including 3D organoids and organ-on-a-chip technologies provide more translationally relevant platforms for this validation [104].
Emerging Opportunities
Future advancements in this field will likely focus on:
Temporal Modeling: Incorporating longitudinal data to capture dynamic disease processes rather than static snapshots [101].
Personalized Therapeutic Screening: Using patient-specific iPSC models combined with AI-driven analysis to predict individual drug responses and optimize treatment strategies [106] [104].
Spatial Multi-Omics: Integrating spatial transcriptomics and proteomics with cellular phenotypic data to preserve architectural context in AD brains [101].
The integration of AI-driven multi-omics approaches with iPSC-based disease models represents a paradigm shift in Alzheimer's disease research. By implementing the methodologies and frameworks outlined in this technical guide, researchers can leverage these advanced technologies to unravel the complexity of AD mechanisms, identify clinically relevant biomarkers, and accelerate the development of effective therapeutics.
Conclusion
iPSC technology has unequivocally established itself as a transformative force in Alzheimer's disease research, providing unprecedented access to patient-specific human neurons for mechanistic studies and drug discovery. These models successfully recapitulate key pathological features, from Aβ and tau pathology to neuroinflammation and synaptic dysfunction, offering a more human-relevant platform than traditional models. The successful identification of drug combinations and the initiation of clinical trials based on iPSC findings mark a significant leap toward translational impact. Future progress hinges on standardizing protocols, improving the modeling of aging and complex neural circuits, and fully integrating multi-omics and AI-driven approaches. The continued evolution of iPSC-based models promises to accelerate the development of personalized, effective therapeutics for Alzheimer's disease, ultimately bridging the long-standing gap between preclinical discovery and clinical success.
References
- 1. 2025 NIH Alzheimer's Disease and Related Dementias ... [https://www.nia.nih.gov/about/2025-nih-dementia-research-progress-report]
- 2. Modeling Alzheimer’s disease with human iPS cells: advancements, lessons, and applications - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC6689423/]
- 3. Modeling Alzheimer’s disease with iPSC-derived brain cells - PMC [https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6906186/]
- 4. Alzheimer’s in a dish – induced pluripotent stem cell-based disease modeling - PMC [https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6624934/]
- 5. Amyloid Plaque and Neurofibrillary Tangle Pathology in a ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC2527075/]
- 6. Neurons derived from sporadic Alzheimer’s disease iPSCs reveal elevated TAU hyperphosphorylation, increased amyloid levels, and GSK3B activation | Alzheimer's Research & Therapy | Full Text [https://alzres.biomedcentral.com/articles/10.1186/s13195-017-0317-z]
- 7. Exposure of iPSC-derived human microglia to brain ... [https://www.nature.com/articles/s41590-023-01558-2]
- 8. Spatiotemporal analysis of 3D human iPSC-derived neural ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10679684/]
- 9. Drug discovery research with the iPSC models of ... [https://www.sciencedirect.com/science/article/pii/S0168010225001683]
- 10. The Genetics of Alzheimer Disease - PMC - PubMed Central [https://pmc.ncbi.nlm.nih.gov/articles/PMC3475404/]
- 11. Alzheimer's disease patient brain extracts induce multiple ... [https://www.nature.com/articles/s41380-025-03041-w]
- 12. Human induced pluripotent stem cell models for ... [https://www.frontiersin.org/journals/human-neuroscience/articles/10.3389/fnhum.2025.1548701/full]
- 13. Modeling common Alzheimer's disease with high and low ... [https://www.sciencedirect.com/science/article/pii/S2213671125001742]
- 14. Identification of 16 novel Alzheimer's disease loci using multi ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11852348/]
- 15. CRISPR/Cas9 and iPSC-Based Therapeutic Approaches ... [https://www.mdpi.com/2076-3921/14/7/781]
- 16. Validation of a functional human AD model with four ... [https://www.nature.com/articles/s41598-024-73869-9]
- 17. Newly discovered genetic marker could pave the way for ... [https://medicine.iu.edu/news/2024/10/new-genetic-marker-could-pave-way-future-alzheimers-disease-therapeutics]
- 18. Modeling Alzheimer's disease with iPSC-derived brain cells [https://www.nature.com/articles/s41380-019-0468-3]
- 19. Human iPSC-derived glia models for the study of ... - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC10566197/]
- 20. CRISPR/Cas9 and iPSC-Based Therapeutic Approaches in ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12291668/]
- 21. Changes in iPSC-astrocyte morphology reflect Alzheimer's ... [https://pubmed.ncbi.nlm.nih.gov/39704342/]
- 22. Astrocyte and oligodendrocyte pathology in Alzheimer's ... [https://www.sciencedirect.com/science/article/pii/S1878747925000182]
- 23. Human iPSC-Derived Neural Models for Studying Alzheimer's ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8930932/]
- 24. Harnessing human iPSC-microglia for CNS-wide delivery ... [https://www.sciencedirect.com/science/article/pii/S1934590925000992]
- 25. Human Induced Pluripotent Stem Cell-based Disease ... [https://www.sciencedirect.com/science/article/pii/S0306452222002354]
- 26. Alzheimer's in a dish – induced pluripotent stem cell-based ... [https://translationalneurodegeneration.biomedcentral.com/articles/10.1186/s40035-019-0161-0]
- 27. Alzheimer's disease and synapse Loss: What can we learn ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10703628/]
- 28. Subcellular proteomics and iPSC modeling uncover ... [https://www.nature.com/articles/s43587-025-00823-3]
- 29. Large-scale drug screening in iPSC-derived motor neurons ... [https://www.nature.com/articles/s41593-025-02118-7]
- 30. Modeling common Alzheimer's disease with high and low ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12365843/]
- 31. The APOE isoforms differentially shape the transcriptomic and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12106835/]
- 32. A 3D human iPSC-derived multi-cell type neurosphere system ... [https://jneuroinflammation.biomedcentral.com/articles/10.1186/s12974-025-03433-3]
- 33. Drug discovery research with the iPSC models of ... [https://pubmed.ncbi.nlm.nih.gov/41213329/]
- 34. Induced pluripotent stem cells (iPSCs) [https://www.nature.com/articles/s41392-024-01809-0]
- 35. Differentiation of human induced pluripotent stem cells into cortical neural stem cells - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC9849742/]
- 36. A functional hiPSC-cortical neuron differentiation and maturation model and its application to neurological disorders - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC8758945/]
- 37. A simplified protocol for differentiation of electrophysiologically mature neuronal networks from human induced pluripotent stem cells | Molecular Psychiatry [https://www.nature.com/articles/mp201756]
- 38. Progress and challenges in directing the differentiation of ... [https://www.frontiersin.org/journals/cell-and-developmental-biology/articles/10.3389/fcell.2022.1089970/full]
- 39. Large-scale differentiation of iPSC-derived motor neurons ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10557526/]
- 40. Induced Pluripotent Stem Cells in Drug Discovery and ... [https://pubmed.ncbi.nlm.nih.gov/38397069/]
- 41. Human 3D brain organoids: steering the ... - Nature [https://www.nature.com/articles/s41420-023-01523-w]
- 42. From 2D to 3D Co-Culture Systems - PubMed Central - NIH [https://pmc.ncbi.nlm.nih.gov/articles/PMC9656609/]
- 43. A review of protocols for brain organoids and applications for ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9803834/]
- 44. Visualization of morphological categories of colonies for ... [https://www.sciencedirect.com/science/article/pii/S235232041630058X]
- 45. Recent Advances in High-Content Imaging and Analysis in ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10572296/]
- 46. Organoids, 3D Organoid Research [https://www.moleculardevices.com/applications/3d-cell-models/organoids]
- 47. High-volume, label-free imaging for quantifying single-cell ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10878516/]
- 48. Development of a platform to investigate long-term ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9481914/]
- 49. Induced long-term potentiation improves synaptic stability ... [https://www.sciencedirect.com/science/article/pii/S0969996125002943]
- 50. Human Pluripotent Stem Cell-Derived Models and Drug ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8193821/]
- 51. An Electrophysiological and Pharmacological Study of the Properties of... [https://pubmed.ncbi.nlm.nih.gov/34440722/]
- 52. High-content imaging as a tool to quantify and characterize ... [https://www.sciencedirect.com/science/article/pii/S2667237523001455]
- 53. 332. Examples of scientific data visualization with ... [https://medium.com/@ilakk2023/examples-of-scientific-data-visualization-with-quantitative-components-77ec53b7ce43]
- 54. 80 types of charts & graphs for data visualization (with ... [https://www.datylon.com/blog/types-of-charts-graphs-examples-data-visualization]
- 55. Network-Medicine-Guided Drug Repurposing for Alzheimer's ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12563243/]
- 56. Network-Medicine-Guided Drug Repurposing for ... [https://www.mdpi.com/1422-0067/26/20/10003]
- 57. Drug repurposing for Alzheimer's disease and other ... [https://www.nature.com/articles/s41467-025-56690-4]
- 58. The role for artificial intelligence in identifying combination ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12627895/]
- 59. Addressing variability in iPSC-derived models of human ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6994963/]
- 60. Analysis of transcriptional variability in a large human iPSC ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5384872/]
- 61. Systematic assessment of variability in the proteome ... [https://www.sciencedirect.com/science/article/pii/S1873506121003597]
- 62. Improved modeling of human AD with an automated ... [https://www.nature.com/articles/s41467-021-25344-6]
- 63. Back and Forth in Time: Directing Age in iPSC-Derived Lineages [https://pmc.ncbi.nlm.nih.gov/articles/PMC4870156/]
- 64. Strategies for modeling aging and age-related diseases [https://www.nature.com/articles/s41514-024-00161-5]
- 65. Current Development of iPSC-Based Modeling in ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12027653/]
- 66. Large-scale generation of human iPSC-derived neural stem ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4594592/]
- 67. Potential use of iPSCs for disease , drug screening, and... modeling [https://pmc.ncbi.nlm.nih.gov/articles/PMC10687886/]
- 68. A high efficiency precision genome editing method with ... [https://www.nature.com/articles/s41598-024-60766-4]
- 69. CRISPR-based functional genomics for neurological disease [https://pmc.ncbi.nlm.nih.gov/articles/PMC7484261/]
- 70. New CRISPR/Gene Editing Strategies for Alzheimer's [https://aaic.alz.org/releases_2023/crispr-gene-editing-treatment-potential-alzheimers.asp]
- 71. Subcellular proteomics and iPSC uncover... | Nature Aging modeling [https://www.nature.com/articles/s43587-025-00823-3?error=cookies_not_supported]
- 72. BRAIN 2025: A Scientific Vision - BRAIN Initiative - NIH [http://braininitiative.nih.gov/vision/nih-brain-initiative-reports/brain-2025-scientific-vision]
- 73. Modeling Alzheimer's disease with iPSC-derived brain cells [https://pmc.ncbi.nlm.nih.gov/articles/PMC6906186/]
- 74. Open Data In Neurophysiology: Advancements, Solutions ... [https://www.eneuro.org/content/12/11/ENEURO.0486-24.2025]
- 75. Top 9 Types of Charts in Data Visualization [https://365datascience.com/trending/chart-types-and-how-to-select-the-right-one/]
- 76. Understanding WCAG 2 Contrast and Color Requirements [https://webaim.org/articles/contrast/]
- 77. Human induced pluripotent stem cell models for Alzheimer's ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11962003/]
- 78. Modeling Alzheimer's disease with human induced pluripotent ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5930170/]
- 79. A comparative review of cell culture systems for the study of ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3407712/]
- 80. Cell models for Alzheimer's and Parkinson's disease [https://www.sciencedirect.com/science/article/pii/S0753332222003134]
- 81. The Challenges to Advancing Induced Pluripotent Stem ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10768945/]
- 82. Exploring iPSC-Derived Cells in Alzheimer's Research [https://communities.springernature.com/posts/exploring-ipsc-derived-cells-in-alzheimer-s-research-from-mouse-models-to-human-insights]
- 83. Current Challenges of iPSC-Based Disease Modeling and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6562607/]
- 84. A lightweight machine learning tool for Alzheimer's disease ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12620993/]
- 85. Machine learning in Alzheimer's disease genetics [https://www.nature.com/articles/s41467-025-61650-z]
- 86. Potential use of iPSCs for disease modeling, drug screening, and cell-based therapy for Alzheimer’s disease | Cellular & Molecular Biology Letters | Full Text [https://cmbl.biomedcentral.com/articles/10.1186/s11658-023-00504-2]
- 87. Postmortem Brain Imaging in Alzheimer's Disease and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10693476/]
- 88. Single-cell multiregion dissection of Alzheimer's disease [https://www.nature.com/articles/s41586-024-07606-7]
- 89. Large-scale study of brain proteins uncovers new clues to ... [https://www.alzheimers.gov/news/large-scale-study-brain-proteins-uncovers-new-clues-alzheimers-disease]
- 90. The Amyloid-β Pathway in Alzheimer's Disease [https://www.nature.com/articles/s41380-021-01249-0]
- 91. Pathophysiological subtypes of Alzheimer's disease based ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7805814/]
- 92. 综述:利用人iPSCs建立神经退行性疾病治疗方法 [https://www.ebiotrade.com/newsf/2025-10/20251031084903189.htm]
- 93. Human iPSC -Derived Neural Models for Studying... [https://link.springer.com/article/10.1007/s12015-021-10254-3]
- 94. Modeling Alzheimer's Disease with Induced Pluripotent ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4895035/]
- 95. Modeling Alzheimer ’ s with disease -derived brain cells iPSC [https://www.nature.com/articles/s41380-019-0468-3?error=cookies_not_supported&code=6baed35a-b1ae-4f33-badf-c1e925b35134]
- 96. 一种功能性的老年人类iPSC-皮质神经元模型揭示AD机制 [https://www.brainmed.com/info/detail?id=48211]
- 97. Ropinirole, a New ALS Drug Candidate Developed Using ... [https://www.sciencedirect.com/science/article/pii/S0165614719302743]
- 98. Artificial intelligence-driven multi-omics approaches in ... [https://www.sciencedirect.com/science/article/pii/S2211383525005088]
- 99. AI-assisted multi-OMICS analysis reveals new markers for ... [https://pubmed.ncbi.nlm.nih.gov/40441366/]
- 100. PhenoModel: A multimodal phenotypic drug design ... [https://www.sciencedirect.com/science/article/pii/S2211383525006446]
- 101. Decoding neuroinflammation in Alzheimer's disease [https://www.frontiersin.org/journals/immunology/articles/10.3389/fimmu.2025.1616899/full]
- 102. Modeling common Alzheimer's disease with high and low ... [https://pubmed.ncbi.nlm.nih.gov/40614728/]
- 103. iPSCs from the Alzheimer's Disease Neuroimaging ... [https://card.nih.gov/research-programs/ida-project]
- 104. AI in Phenotypic Drug Discovery & Research [https://lifesciences.danaher.com/us/en/library/ai-in-phenotypic-drug-discovery.html]
- 105. From Phenotypic Screening to AI-Powered Insight [https://ardigen.com/from-phenotypic-screening-to-ai-powered-insight-how-to-generate-data-to-get-the-most-from-ardigen-phenaid/]
- 106. Artificial Intelligence (AI) Applications in Drug Discovery and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11510778/]