Modeling Neurodevelopmental Disorders with Human iPSCs: From Foundational Mechanisms to Translational Applications
This article provides a comprehensive overview of how human induced pluripotent stem cells (iPSCs) are revolutionizing the study of neurodevelopmental disorders (NDDs).
Modeling Neurodevelopmental Disorders with Human iPSCs: From Foundational Mechanisms to Translational Applications
Abstract
This article provides a comprehensive overview of how human induced pluripotent stem cells (iPSCs) are revolutionizing the study of neurodevelopmental disorders (NDDs). Covering foundational principles to cutting-edge applications, we explore the transition from traditional 2D models to complex 3D organoid and assembloid systems that better recapitulate human brain development and disease phenotypes. The content details advanced methodological approaches, including CRISPR genome editing, functional neuronal assays, and machine learning integration, while addressing key challenges in standardization, scalability, and clinical translation. Aimed at researchers, scientists, and drug development professionals, this review synthesizes how iPSC-based models are accelerating disease mechanism discovery and creating new pathways for therapeutic development in conditions affecting the developing human nervous system.
The iPSC Revolution: Establishing Human-Centric Models for Neurodevelopmental Disorders
The study of neurodevelopmental and neurodegenerative disorders has long relied on animal models. While these models have provided foundational knowledge, they fundamentally lack human-specific biological context, often leading to findings that fail to translate to human patients. Animal models face significant challenges in recapitulating human brain complexity due to evolutionary divergence in brain structure, function, and genetics [1]. For instance, key parameters such as heart rates differ dramatically between species (zebrafish: 120-180 bpm; mice: 300-600 bpm; humans: 60-100 bpm), reflecting profound underlying physiological differences that complicate direct translation of neurological findings [1]. Furthermore, species-specific disease mechanisms often mean that pathologies observed in animals do not fully mirror human disease states, particularly for complex neurodevelopmental disorders like schizophrenia or autism spectrum disorders [2].
The emergence of induced pluripotent stem cell (iPSC) technology has revolutionized neurological disease modeling by providing access to functional human neural cells and systems. iPSCs are particularly valuable for neural research because they can be derived from patients with known symptom histories, genetics, and drug-response profiles, capturing the full genetic complexity of polygenic brain disorders in a human-relevant system [2]. This approach enables researchers to move beyond the limitations of animal models to create human neural systems that authentically replicate disease mechanisms and developmental processes.
The iPSC Revolution in Neural System Modeling
Fundamental Principles and Advantages
The development of iPSC technology by Takahashi and Yamanaka in 2006-2007 represented a paradigm shift in disease modeling [3]. By reprogramming somatic cells to a pluripotent state using defined factors (OCT4, SOX2, KLF4, MYC), researchers gained the ability to generate patient-specific neural cells that retain the complete genetic background of the donor [3]. This technology provides three critical advantages for neural research: (1) the capacity for unlimited expansion of patient-specific neural progenitors, (2) amenability to genetic engineering using CRISPR/Cas9 to create isogenic controls, and (3) the ability to differentiate into diverse neural and glial subtypes relevant to specific disorders [2] [3].
iPSC-derived neural models effectively capture human-specific aspects of brain development that cannot be studied in animal models. Notably, human brain development features a protracted period of neurogenesis and interneuron migration that extends into the postnatal period—a uniquely human characteristic with profound implications for neurodevelopmental disorders [4]. Recent findings in postmortem tissue have revealed that cortical interneuron migration continues after birth, "shed[ding] light on a prolonged stage of human brain development and a longer plasticity window for fine-tuning of the developing circuit with local inhibitory inputs" [4]. This extended developmental window also presents a wider vulnerability to insults that may lead to neurological disorders such as autism and epilepsy [4].
Technical Validation of iPSC-Derived Neural Models
Extensive validation studies have confirmed that iPSC-derived neural cells accurately model human neural development. Molecular marker analyses, morphological assessments, and functional assays demonstrate that iPSC-derived neurons exhibit characteristics of authentic human neural cells [2]. Transcriptome-wide approaches using single-cell RNA sequencing have enabled high-dimensional, unbiased validation of iPSC-derived cellular models, confirming they give rise to heterogeneous, region-specific cell types [2].
Temporal analyses consistently show that iPSC-derived neurons correspond to prenatal developmental stages, most commonly resembling the second trimester of human gestation [2]. This developmental timing makes them particularly suitable for studying neurodevelopmental disorders that originate during early brain development. While this fetal-like state presents challenges for modeling late-onset disorders, recent advances in prolonged culture systems and maturation protocols have extended the developmental timeline accessible to researchers [4].
Table 1: Quantitative Comparison of Neural Model Systems
| Model Characteristic | Animal Models | iPSC 2D Models | iPSC 3D Organoids/Assembloids |
|---|---|---|---|
| Human genetic background | No (unless humanized) | Yes | Yes |
| Developmental stage captured | Species-specific | ~Second trimester human equivalent | Multiple developmental stages |
| Cellular diversity | Species-specific repertoire | Limited by protocol | High diversity, self-organizing |
| Circuit complexity | Intact but non-human | Limited connections | Emerging complex connectivity |
| Neuroinflammatory components | Limited human relevance | Limited | Advanced (microglia, astrocytes) |
| Throughput for screening | Low | High | Medium |
| Maturation timeline | Fixed developmental program | 30-90 days | Up to 390+ days [4] |
Advanced Human Neural System Architectures
From 2D Cultures to 3D Organoids and Assembloids
Early iPSC neural models primarily consisted of two-dimensional monocultures of specific neural subtypes. While these provided important insights into cell-autonomous disease mechanisms, the field has progressively advanced toward more complex three-dimensional systems that better replicate the cellular diversity and spatial organization of the human brain [3]. Cerebral organoids—self-organizing 3D structures that contain multiple neural cell types—represent a significant advancement, exhibiting features of regional specification and layered organization reminiscent of the developing human brain [3].
The most recent innovation involves assembloids—integrated systems combining multiple organoid types to model interactions between different brain regions. One groundbreaking study established a dorsal-ventral assembloid model that reconstitutes postnatal human interneuron migration [4]. This model, maintained for up to 390 days in culture, demonstrated that "newly born migratory interneurons arrange themselves into connected chains that are surrounded by astrocytes," replicating the architectural and migratory patterns observed in early postnatal human brains [4]. Electron microscopy analysis revealed "architecture essentially indistinguishable from what has been seen in early postnatal human brains," validating this approach for studying extended human brain development [4].
Incorporation of Neuroimmune Components
The critical role of neuroinflammation in neurodegenerative diseases has driven the development of iPSC models that incorporate diverse glial cell types. Recent advancements now enable the differentiation of iPSCs into microglia, astrocytes, and blood-brain barrier (BBB) components, which can be integrated into complex organoid systems [5]. These advancements are particularly relevant for modeling neuroinflammation in prevalent neurodegenerative disorders such as Alzheimer's disease (AD), Parkinson's disease (PD), and multiple sclerosis (MS) [5].
The integration of neuroimmune components enables researchers to study cell-non-autonomous disease mechanisms, such as how microglial activation contributes to neuronal damage in neurodegeneration, or how astrocyte dysfunction impacts synaptic pruning in neurodevelopmental disorders. These human-specific neuroimmune interactions cannot be adequately modeled in animal systems due to significant species differences in immune function and inflammation responses [5].
Application to Neurodevelopmental Disorder Research
Modeling Genetic Complexity in Schizophrenia
Schizophrenia (SCZ) research exemplifies the power of iPSC systems to address polygenic complexity. Despite high heritability, genome-wide association studies have identified numerous risk variants with low penetrance, making it difficult to establish causal relationships [2]. iPSC models capture the complete genetic background of patients, enabling researchers to study how multiple genetic variants interact to produce disease phenotypes.
A recent innovative approach addressed this complexity through "village editing"—CRISPR/Cas9 gene editing in a cell village format [4]. Researchers generated NRXN1 knockouts in iPSC lines from 15 donors with low, neutral, or high polygenic risk scores for SCZ, achieving high editing efficiency (heterozygous: 33.1%; homozygous: 28.4%) [4]. After differentiation into cortical excitatory neurons, transcriptomic analysis revealed that "genetic background deeply influences gene expression changes in NRXN1 KO neurons" [4]. This demonstrates the critical importance of incorporating multiple genetic backgrounds when studying polygenic disorders and provides a framework for efficient development of similar tools for other complex disorders.
Elucidating Disease Mechanisms in Rare Genetic Disorders
For monogenic neurodevelopmental disorders, iPSC models enable precise dissection of disease mechanisms. In Hereditary Sensory and Autonomic Neuropathy Type IV (HSAN IV), caused by mutations in NTRK1, iPSC-derived dorsal root ganglion (DRG) organoids revealed a previously unknown disease mechanism: "DRG organoids derived from HSAN IV patients underwent a lineage switching between sensory neurons and glial cells without affecting the neural crest stem cell population" [4]. This lineage switching, characterized by reduced sensory neurons and premature gliogenesis, represents a novel pathological mechanism that could not have been identified in animal models [4].
The use of isogenic controls—patient-derived iPSCs in which the disease-causing mutation has been corrected using CRISPR—eliminates confounding genetic variability and provides definitive evidence of mutation-specific effects [4]. This approach is particularly powerful for validating potential therapeutic targets by demonstrating that phenotype rescue occurs specifically upon mutation correction.
Table 2: Key Research Reagent Solutions for iPSC Neural Disease Modeling
| Research Reagent/Tool | Function/Application | Example in Neurodevelopmental Research |
|---|---|---|
| Sendai virus, episomal plasmids, mRNA | Non-integrating reprogramming methods | Generating integration-free iPSCs from patient fibroblasts or blood cells [2] |
| CRISPR/Cas9 gene editing | Creating isogenic controls; introducing mutations | NRXN1 knockout in multiple genetic backgrounds [4]; correction of NTRK1 mutations [4] |
| Neural patterning molecules | Directing differentiation to specific neural fates | Generating cortical excitatory neurons, GABAergic interneurons, etc. [2] |
| OP9 stromal cells | Supporting hematopoietic differentiation | Co-culture system for iPSC differentiation [6] |
| Tandem Mass Tag (TMT) labeling | Multiplexed quantitative proteomics | Comparing proteomes of iPSC-derived cells vs. primary cells [6] |
| Single-cell RNA sequencing | High-resolution cell type characterization | Validating neuronal subtypes; identifying novel populations [2] |
Methodological Framework for iPSC Neural System Experiments
Experimental Workflow for Disease Modeling
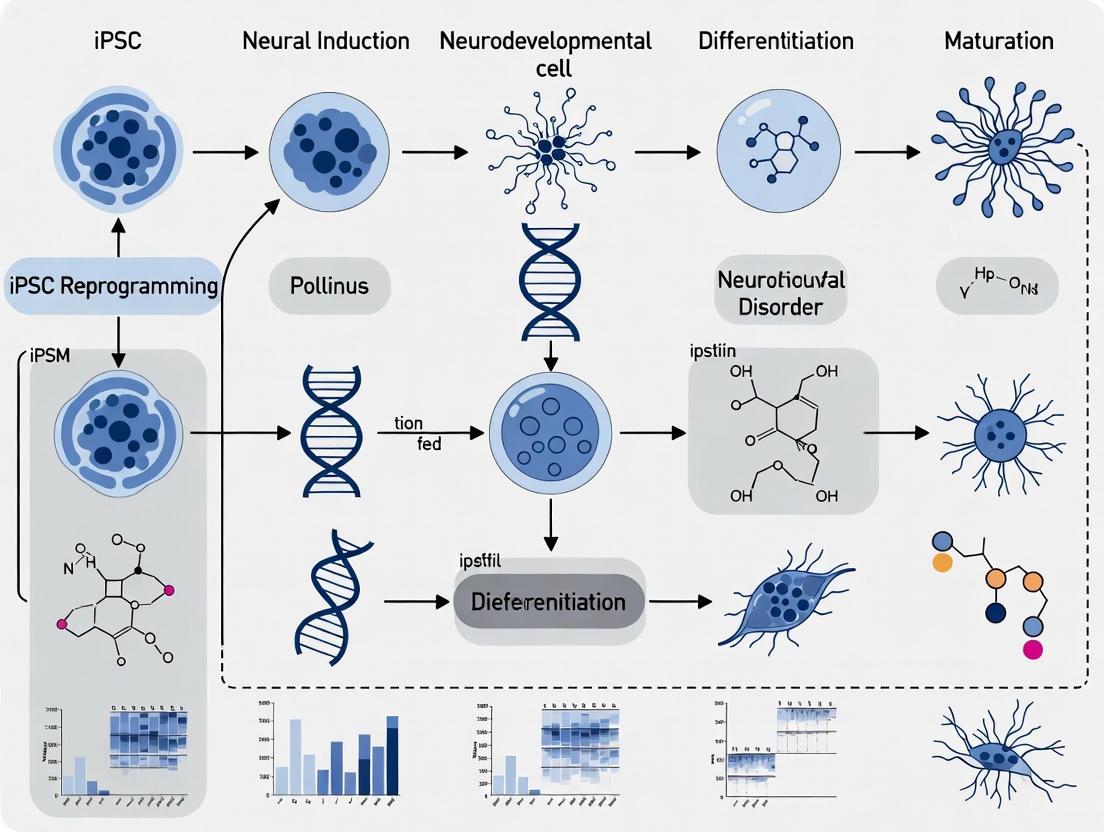
The following diagram illustrates a comprehensive workflow for iPSC-based modeling of neurodevelopmental disorders:
Protocol for Dorsal-Ventral Assembloid Generation
The assembloid model of interneuron migration represents a cutting-edge approach for studying human-specific developmental processes [4]. The detailed methodology includes:
Independent Differentiation: Dorsal (cortical) and ventral (ganglionic eminence) organoids are differentiated separately from iPSCs using established patterning protocols with appropriate morphogens.
Fusion Protocol: After 120 days of differentiation, dorsal and ventral organoids are placed in contact in low-adhesion plates to enable fusion.
Long-term Maintenance: Fused assembloids are maintained for extended periods (up to 390 days) with careful feeding schedules and periodic assessment.
Migration Analysis: Interneuron migration from ventral to dorsal compartments is assessed using time-lapse imaging, immunohistochemistry for CGE markers, and EdU birth dating to confirm late-born interneuron populations.
Validation: Electron microscopy and single-cell spatial transcriptomics validate architectural features and molecular signatures comparable to postnatal human brain tissue [4].
This protocol successfully reconstitutes "events of human brain development that occur after birth, allowing a genetic and cell biological analysis of this important phenomenon" [4].
Protocol for Village Editing Approach
The village editing approach enables efficient study of genetic variants across multiple backgrounds [4]:
iPSC Line Selection: Select iPSC lines from multiple donors (e.g., 15 donors) representing diverse polygenic risk backgrounds.
Pooled CRISPR Editing: Perform CRISPR/Cas9 editing (e.g., NRXN1 knockout) on a pooled "village" of iPSCs from multiple donors rather than editing lines individually.
Clonal Isolation and Genotyping: After editing, recover individual clones and genotype to identify untargeted controls, heterozygous (33.1%), and homozygous (28.4%) edits across different donor backgrounds.
Differentiation and Analysis: Differentiate edited iPSCs into relevant neural cells (e.g., cortical excitatory neurons) and analyze transcriptomic, synaptic, or other cellular phenotypes.
Background Effect Assessment: Use computational methods to determine how genetic background influences the phenotypic effects of the introduced mutation [4].
This method provides "a framework for rapid and efficient development of similar tools to study gene functions in complex, polygenic disorders" [4].
Quantitative Assessment of Model System Faithfulness
Rigorous comparison between iPSC-derived neural cells and their in vivo counterparts is essential for validating model systems. Multiplexed quantitative proteomics using Tandem Mass Tag (TMT) labeling has demonstrated that iPSC-derived erythroid cells share significant similarity with primary cells, with only 1.9% of proteins differing by 5-fold or more [6]. While direct proteomic comparisons for neural cells were not provided in the search results, similar validation approaches are being applied to neural lineages.
Table 3: Functional Comparison of Neural Model Capabilities
| Model Capability | Traditional Animal Models | iPSC 2D Neural Cultures | iPSC 3D Organoid/Assembloid Systems |
|---|---|---|---|
| Globin switching study | Limited relevance | Not applicable | Not applicable |
| Enucleation analysis | Species-specific process | Not applicable | Not applicable |
| Neural migration studies | Limited human relevance | Limited | High (chain migration observed) [4] |
| Lineage specification analysis | Requires transgenic models | Moderate | High (lineage switching detected) [4] |
| Cell-cell interaction mapping | Intact but non-human | Limited | High (neuron-astrocyte interactions) [4] |
| Drug screening throughput | Low | High | Medium |
| Transcriptomic profiling | Species-specific | Human-specific | Human-specific, spatially resolved |
Future Directions and Integration with Emerging Technologies
The next frontier in human neural system development involves integrating iPSC models with advanced computational approaches and additional technological innovations. Machine learning is emerging as a powerful tool for analyzing complex datasets generated from iPSC models and for predicting disease outcomes based on in vitro phenotypes [1]. As these models become more sophisticated, they will increasingly incorporate multi-omics approaches (transcriptomics, proteomics, epigenomics, metabolomics) to build comprehensive pictures of disease states.
Another promising direction is the development of humanized animal models through transplantation of human iPSC-derived neural cells or organoids into rodent brains, creating chimeric systems that combine the physiological context of animal models with human cellular components [1]. These approaches may provide important bridges for translational research while maintaining human biological relevance.
Furthermore, the combination of iPSC-derived neural models with advanced bioengineering approaches—such as microfluidic devices, bioprinting, and electrical stimulation—will enable even more precise control over the cellular microenvironment and more complex tissue architectures that better mimic the human brain [3].
Human iPSC-derived neural systems represent a transformative approach for studying neurodevelopmental disorders, offering unprecedented access to human-specific biology and disease mechanisms. By capturing the complete genetic background of patients and enabling the reconstruction of complex neural tissues in vitro, these models address critical limitations of traditional animal systems. The continued refinement of organoid and assembloid technologies, combined with advanced gene editing and multi-omics characterization, promises to accelerate our understanding of neurodevelopmental disease mechanisms and therapeutic development. As these human-relevant systems become increasingly sophisticated and accessible, they will undoubtedly play a central role in unraveling the complexities of human brain development and dysfunction.
The modeling of neurodevelopmental disorders (NDDs) has been fundamentally transformed by technologies that enable the reprogramming of somatic cells into specific neural lineages [7]. For decades, neuroscience research was constrained by the limited accessibility of human neural tissue and the imperfect translatability of animal models, which fail to fully capture the intricacies of human-specific developmental processes [7]. The emergence of human induced pluripotent stem cells (hiPSCs) inaugurated a new era, allowing researchers to derive patient-specific neurons, glia, and three-dimensional (3D) organoid systems that more accurately model human physiology and pathology [7]. These advances are particularly crucial for NDDs—a highly heterogeneous group of diseases impairing social, cognitive, and emotional functioning—as they provide direct experimental access to disease mechanisms within a human genetic context [7]. This technical guide outlines the core principles, methods, and applications of somatic cell reprogramming and neural specification, providing a comprehensive framework for researchers investigating NDD mechanisms.
Somatic Cell Reprogramming: Methodologies and Mechanisms
Historical Foundations and Key Discoveries
The conceptual foundation for somatic cell reprogramming was established by pioneering work demonstrating that cellular differentiation is not an irreversible process. John Gurdon's seminal somatic cell nuclear transfer (SCNT) experiments in Xenopus laevis in 1962 first revealed that a nucleus from a terminally differentiated somatic cell contained all genetic information needed to generate an entire organism [3]. This principle of epigenetic plasticity was later harnessed by Shinya Yamanaka, who identified a combination of four transcription factors—OCT4, SOX2, KLF4, and MYC (OSKM)—sufficient to reprogram mouse fibroblasts into induced pluripotent stem cells (iPSCs) in 2006 [3]. This discovery, followed by the successful generation of human iPSCs in 2007, established the core technology that enables the current modeling of human diseases, including NDDs [3].
Reprogramming Strategies and Technical Approaches
Multiple strategic pathways exist for converting somatic cells to neural lineages, each with distinct advantages for specific research applications. The table below summarizes the primary reprogramming approaches.
Table 1: Strategic Pathways for Neural Reprogramming
| Reprogramming Strategy | Key Features | Intermediate Stage | Technical Considerations | Primary Research Applications |
|---|---|---|---|---|
| Induced Pluripotency [8] [3] | Somatic cells are fully reprogrammed to a pluripotent state using Yamanaka factors (OSKM) or related combinations. | Induced Pluripotent Stem Cells (iPSCs) | Lengthy process (months); potential for teratoma formation; captures complete developmental trajectory. | Disease modeling requiring full neurodevelopment; generation of diverse neural cell types; organoid formation. |
| Direct Lineage Reprogramming (Transdifferentiation) [8] [9] | Somatic cells are directly converted to neuronal cells using neural-specific transcription factors or small molecules, bypassing the pluripotent state. | None | Faster process (weeks); reduced tumorigenic risk; may result in incomplete maturation. | Rapid generation of specific neuronal subtypes; potential for in vivo therapeutic applications. |
| Induced Neural Stem Cells (iNSCs) [8] | Somatic cells are reprogrammed into multipotent neural stem cells capable of self-renewal and differentiation into multiple neural lineages. | Neural Stem Cells (NSCs) | Stable, expandable cell population; maintains lineage restriction; differentiates into neurons, astrocytes, and oligodendrocytes. | Studies requiring expandable progenitor populations; modeling early neurodevelopmental events. |
Molecular Mechanisms of Reprogramming
The reprogramming of somatic cells to iPSCs involves profound epigenetic remodeling that partially reverses the process of embryonic development [3]. This process occurs in two broad phases: an early, stochastic phase where somatic genes are silenced and early pluripotency genes are activated, and a late, more deterministic phase where late pluripotency-associated genes are activated [3]. Critical events during reprogramming include widespread changes in chromatin accessibility, DNA methylation patterns, and histone modifications [3]. When reprogramming fibroblasts, a key event is the mesenchymal-to-epithelial transition (MET), which is crucial for establishing the pluripotent state [3]. The entire process entails a comprehensive resetting of the cellular state, affecting nearly all aspects of cell biology, including metabolism, cell signaling, and proteostasis [3].
Neural Lineage Specification: From Pluripotency to Neuronal Networks
Guided Neural Induction and Regional Patterning
The controlled differentiation of iPSCs into neural lineages requires precise manipulation of key developmental signaling pathways. Highly efficient neural induction can be achieved using small-molecule cocktails that suppress alternative differentiation paths while promoting neural specification [10]. One established protocol involves a "DAP" cocktail (typically containing Dorsomorphin, A83-01, and other factors) that inhibits SMAD signaling, leading to highly pure cultures of PAX6-/NESTIN-positive neural stem cells (NSCs) with greater than 97% efficiency [10]. These primitive NSCs can subsequently be patterned into specific neuronal subtypes through the sequential addition of regionalizing factors:
- Cortical Excitatory Neurons: Treatment with FGF2 and WNT antagonists promotes anterior forebrain identities, while timed WNT activation can drive posteriorization [10] [4].
- Interneuron Subtypes: Activation of SHH signaling ventralizes neural progenitor cells towards medial ganglionic eminence identities that generate GABAergic interneurons [4].
- Sensory Neurons: Combined BMP, WNT, and TGF-β signaling directs differentiation towards neural crest lineages that form sensory neurons, as demonstrated in dorsal root ganglion (DRG) organoid models [4].
Advanced 3D Model Systems: Organoids and Assembloids
Beyond two-dimensional cultures, 3D brain organoids recapitulate more complex features of human neurodevelopment, including progenitor proliferation, neuronal migration, and cortical layer formation [7]. These systems are particularly valuable for modeling malformations of cortical development, which often present with microcephaly, disorganized cytoarchitecture, and severe cognitive impairments [7]. Recent advances include the generation of dorsal-ventral assembloids by fusing region-specific organoids, which model the migration of interneurons from ventral to dorsal regions—a process critical for establishing balanced cortical circuitry [4]. These assembloid systems have revealed that late-born interneurons arrange into chain-like structures surrounded by astrocytes, essentially recapitulating postnatal migratory streams observed in human infants [4].
Table 2: Quantitative Proteomic and Phosphoproteomic Landscape During Neural Specification
| Molecular Category | hPSCs | hNSCs | Key Regulated Components | Functional Significance |
|---|---|---|---|---|
| Total Proteins Identified [10] | ~13,000 | ~13,000 | Transcription factors, epigenetic regulators | Comprehensive mapping of proteome dynamics |
| Phosphorylation Sites [10] | ~60,000 | ~60,000 | Kinase substrates, signaling nodes | Insight into post-translational regulation |
| Neural Induction Markers | - | - | PAX6, NESTIN, OTX2 | Confirmation of neural lineage commitment |
| Pluripotency Factors | - | - | OCT4, NANOG, SOX2 | Downregulated during specification |
| Validated Regulators | - | - | Midkine (MDK) | Novel secreted factor promoting neuralization |
Applications in Neurodevelopmental Disorder Research
Disease Modeling and Mechanistic Insights
iPSC-derived neural models have become indispensable tools for elucidating the pathophysiology of NDDs. In idiopathic autism spectrum disorder (ASD), iPSC-derived neurons exhibit functional alterations including reduced calcium transients, impaired synaptic neurotransmission, and decreased network connectivity [11]. Molecular profiling of ASD-derived neuronal progenitor cells (NPCs) has identified differentially expressed microRNAs (hsa-let-7e-5p, hsa-miR-135b-5p, hsa-miR-16-5p, and hsa-miR-27b-3p) that cluster in pathways regulating neurogenesis, neuronal functioning, and cAMP/Ca2+ signaling [11]. For monogenic disorders like Tuberous Sclerosis Complex (TSC), iPSC-derived neurons have revealed dysregulation of the mTOR signaling pathway and neuronal hyperexcitability that can be rescued by mTORC1-selective inhibitors [7]. Similarly, iPSC models of epilepsy generated from patients with CLCNKB mutations have uncovered differentially expressed gene networks implicated in epileptogenesis through transcriptomic profiling [7].
High-Content Screening and Therapeutic Development
The integration of iPSC-derived neural models with functional phenotyping platforms enables sophisticated screening applications for NDD research. Multielectrode arrays, calcium imaging, and high-content imaging can capture disease-relevant phenotypes at scale [7]. When combined with machine learning approaches, these rich functional datasets can classify subtle phenotypic signatures, accelerate drug screening, and improve disease modeling in both 2D cultures and 3D organoids [7]. For example, chemogenetic approaches using Designer Receptors Exclusively Activated by Designer Drugs (DREADDs) in co-cultures of human iPSC-derived neurons and rat cortical neurons have demonstrated impaired synaptic connectivity in ASD models [11]. These platforms provide valuable tools for evaluating potential therapeutic compounds in human-relevant systems before advancing to clinical trials.
The Scientist's Toolkit: Essential Reagents and Methodologies
Table 3: Essential Research Reagents for Neural Reprogramming and Differentiation
| Reagent Category | Specific Examples | Function in Protocol | Application Context |
|---|---|---|---|
| Reprogramming Factors | OCT4, SOX2, KLF4, MYC (OSKM) [3]; OCT4, SOX2, NANOG, LIN28 [3] | Induction of pluripotency in somatic cells | iPSC generation from patient fibroblasts or blood cells |
| Neural Induction Molecules | Dorsomorphin, A83-01, SB431542, LDN193189 [10] | SMAD inhibition to promote neural ectoderm formation | Initial neural specification from iPSCs |
| Neural Progenitor Markers | Antibodies against PAX6, NESTIN, SOX1 [10] | Identification and sorting of neural stem cells | Quality control of neural differentiation |
| Neuronal Maturation Factors | BDNF, GDNF, NT-3, cAMP, Ascorbic Acid [11] | Promotion of neuronal survival, maturation, and synaptic development | Terminal differentiation of neurons from NPCs |
| Functional Assay Reagents | GCaMP6s calcium indicator [11], DREADDs [11] | Measurement of neuronal activity and network connectivity | Functional characterization of neuronal models |
Visualizing Signaling Pathways and Workflows
Core Signaling Pathways in Neural Specification
Experimental Workflow for NDD Modeling
The integration of somatic cell reprogramming with neural differentiation technologies has created unprecedented opportunities for modeling neurodevelopmental disorders in a human-specific context. The core principles outlined in this guide—from the initial reprogramming of somatic cells using defined factors to the precise specification of neural lineages through controlled manipulation of developmental signaling pathways—provide a robust framework for investigating disease mechanisms. Current challenges include the standardization of differentiation protocols across laboratories, improvement of functional maturation in iPSC-derived neurons, and meaningful integration of multi-omics datasets [7]. Future advances will likely focus on enhancing the complexity and reproducibility of 3D model systems, incorporating non-neural cell types such as microglia and vascular cells, and developing more sophisticated functional readouts that capture network-level perturbations in NDDs. As these technologies continue to evolve, they promise to bridge the gap between cellular models and clinical applications, ultimately enabling the development of targeted interventions for neurodevelopmental disorders.
The advent of induced pluripotent stem cell (iPSC) technology has ushered in a transformative era for biomedical research, particularly in the study of complex neurodevelopmental disorders. By enabling the reprogramming of adult somatic cells back to a pluripotent state, this technology provides an unprecedented window into human development and disease. The core advantages of patient-specific modeling, preservation of the complete genetic background, and the capacity for unlimited expansion collectively address long-standing limitations of traditional model systems. These properties make iPSCs an indispensable tool for deconstructing the pathogenic mechanisms of neurodevelopmental conditions, which are often influenced by a complex interplay of polygenic and environmental factors that are uniquely human and difficult to recapitulate in animal models [11] [3] [12].
The foundational discovery by Shinya Yamanaka and colleagues, which showed that the forced expression of four transcription factors (Oct4, Sox2, Klf4, and c-Myc) could revert specialized cells to pluripotency, earned them the Nobel Prize in 2012 and established the technical basis for this field [13] [3] [14]. The subsequent refinement of this technology has been driven by the need to create clinically relevant human cellular models that faithfully capture the genetic complexity of individual patients, thereby facilitating a new generation of mechanistic studies and therapeutic discovery efforts [15].
Patient-Specific Modeling of Neurodevelopmental Disorders
Patient-specific modeling stands as a cornerstone advantage of iPSC technology. This approach allows researchers to derive pluripotent stem cells directly from individuals with neurodevelopmental disorders, which can then be differentiated into the specific neural cell types affected in the disease. This process creates a genetically tailored in vitro model system that recapitulates key aspects of the patient's neurobiology.
Applications in Idiopathic and Syndromic Disorders
This capability is particularly powerful for studying idiopathic diseases—those with unknown or complex etiology—where the genetic drivers are not fully understood. For example, in Autism Spectrum Disorder (ASD), a condition characterized by extensive heterogeneity, iPSCs derived from patients with clinically homogeneous presentations have revealed functional alterations in the resulting neurons. These include less frequent calcium transients and impaired synaptic neurotransmission compared to neurons derived from control individuals, providing a measurable cellular phenotype for a behaviorally defined disorder [11]. Similarly, in schizophrenia (SCZ), another polygenic neurodevelopmental condition, iPSC-derived oligodendrocyte precursor cells (OPCs) and oligodendrocytes (OLs) from patients have shown morphological alterations, including significantly increased branch length and junction number in mature oligodendrocytes, pointing to a cell-autonomous, genetically driven deficit in the oligodendroglial lineage that may underlie the white matter disturbances observed in patients [12].
Table 1: Summary of Functional Phenotypes in Patient-Specific iPSC-Derived Neural Cells
| Neurodevelopmental Disorder | iPSC-Derived Cell Type | Key Functional Phenotypes Identified | Citation |
|---|---|---|---|
| Autism Spectrum Disorder (ASD) | Cortical Neurons | ↓ Spontaneous calcium transients; ↓ Synaptic neurotransmission and connectivity; Dysregulated glutamate signaling | [11] |
| Schizophrenia (SCZ) | Oligodendrocytes (OLs) | ↑ Branch length; ↑ Junction number; Dysregulated cell signaling and proliferation | [12] |
Experimental Workflow for Patient-Specific Modeling
The standard workflow for establishing a patient-specific disease model involves several critical steps, each requiring rigorous quality control to ensure the resulting cellular models are accurate and reproducible.
Figure 1: Workflow for patient-specific iPSC modeling of neurodevelopmental disorders.
Preservation of the Native Genetic Background
A paramount strength of iPSC-based models is their ability to preserve the entire and unique genetic blueprint of the donor individual. Unlike other model systems that may introduce confounding genetic variables, iPSCs retain the patient's specific combination of common variants, rare mutations, and structural polymorphisms that collectively contribute to disease susceptibility and manifestation.
Capturing Polygenic Risk in a Dish
Neurodevelopmental disorders like ASD, SCZ, and intellectual disability are highly polygenic. This means that an individual's risk is determined by the combined effect of hundreds or thousands of genetic variants, each with a small individual effect. iPSC models inherently capture this complex genetic architecture. Gene-set enrichment analyses using transcriptomic data from iPSC-derived neural cells have confirmed that the genetic associations identified from large-scale genome-wide association studies (GWAS) for schizophrenia are indeed enriched in the transcriptional signatures of these relevant cell types [12]. This provides a direct biological bridge between statistical genetic findings and functional cellular pathophysiology, allowing researchers to study how a patient's complete set of risk genes orchestrates molecular and functional changes in the brain.
A Platform for Genetic Manipulation
Furthermore, while preserving the native genetic background is crucial for observational studies, the isogenic nature of iPSCs also makes them an ideal platform for targeted genetic manipulation. Using gene-editing tools like CRISPR-Cas9, researchers can introduce or correct specific risk variants in patient-derived iPSC lines. By comparing the edited line to its original parent line, scientists can isolate the phenotypic consequences of a single genetic change against a constant genetic background. This powerful approach allows for the direct demonstration of causality for specific genetic variants identified in patients with neurodevelopmental conditions [13] [3].
Unlimited Expansion Capacity for Research and Screening
The capacity for virtually unlimited self-renewal is a defining property of iPSCs, which has profound practical implications for biomedical research. This characteristic ensures a continuous and scalable supply of biological material, overcoming a major bottleneck that has historically plagued neuroscience: the inability to access living, disease-relevant human brain cells.
Enabling High-Throughput and Reproducible Science
The unlimited expansion potential of iPSCs enables the generation of the large cell numbers required for high-throughput drug screening campaigns, multi-omics studies (genomics, transcriptomics, proteomics), and the development of complex co-culture systems and organoids [13] [3]. This scalability is essential for achieving the statistical power needed to study highly variable human biological systems. Moreover, by creating a master cell bank of a single patient-derived iPSC line, researchers across different labs can perform experiments on a genetically identical, renewable resource, dramatically improving the reproducibility and reliability of findings in neurodevelopmental disease research [16].
Table 2: Applications Enabled by the Unlimited Expansion of iPSCs
| Application | Description | Benefit for Neurodevelopmental Research |
|---|---|---|
| High-Throughput Drug Screening | Testing thousands of small molecules for their ability to reverse cellular phenotypes. | Identifies candidate therapeutics for disorders with no effective medication, e.g., for core symptoms of ASD or SCZ. |
| 'Clinical Trial in a Dish' | Using panels of patient-derived cells to predict variable drug responses. | Personalizes medicine approaches for neurodevelopmental disorders. |
| Multi-omics Profiling | Comprehensive molecular characterization using sequencing and mass spectrometry. | Reveals disease-associated pathways (e.g., synaptic, mTOR, Wnt) from the same biological source. |
| Complex 3D Model Generation | Creating cerebral organoids and assembloids. | Models neural circuit formation and inter-cell-type dysfunction. |
Technical Considerations for Scalable Production
To leverage this advantage for industrial and clinical translation, scalable production processes are being developed. These advanced bioprocessing systems are designed to maintain the integrity of these sensitive cells during large-scale expansion. Studies have demonstrated that integrated systems using low-shear bioreactors and closed, automated cell processing can achieve high cell recovery rates (>90%) and maintain high viability (>95%) and pluripotency marker expression over multiple serial passages, which is critical for generating the quantities of cells needed for robust research and future therapies [16].
The Scientist's Toolkit: Essential Reagents for iPSC-Based Research
The successful application of iPSC technology relies on a suite of specialized reagents and tools, each designed to maintain, differentiate, and characterize pluripotent stem cells and their neuronal progeny.
Table 3: Key Research Reagent Solutions for iPSC Modeling of Neurodevelopmental Disorders
| Reagent/Category | Function | Example Applications |
|---|---|---|
| Reprogramming Factors (OSKM/OSNL) | Core transcription factors (OCT4, SOX2, KLF4, c-MYC or NANOG, LIN28) that initiate reprogramming. | Initial generation of iPSCs from patient somatic cells. |
| Non-Integrating Delivery Vectors | Methods to deliver factors without altering the host genome (e.g., Sendai virus, episomal plasmids, mRNA). | Clinical-grade iPSC generation with enhanced safety profiles. |
| Chemically Defined Media (e.g., mTeSR1, E8) | Standardized, xeno-free nutrient media to support iPSC self-renewal and maintain pluripotency. | Feeder-free culture; maintenance of genomic stability during expansion. |
| Extracellular Matrix Coatings (e.g., Matrigel, Laminin-521) | Surrogate substrate for cell adhesion and signaling, replacing mouse feeder cells. | Feeder-free culture systems that enhance reproducibility. |
| Small Molecule Inhibitors/Activators | Direct differentiation toward specific neural fates (e.g., SMAD, TGF-β, Wnt pathway inhibitors). | Highly efficient generation of cortical neurons, oligodendrocytes, etc. |
| Pluripotency Validation Markers | Antibodies for flow cytometry/immunocytochemistry (e.g., OCT4, SOX2, NANOG, SSEA-4). | Quality control to confirm the pluripotent state of iPSC banks. |
| Genetic Engineering Tools (e.g., CRISPR-Cas9) | For creating isogenic controls or introducing disease-associated mutations. | Establishing causal links between genetic variants and cellular phenotypes. |
Signaling Pathways in Neurodevelopment and Disease
The directed differentiation of iPSCs into specific neural lineages is guided by the sequential activation and inhibition of key evolutionarily conserved signaling pathways. Furthermore, dysregulation of these same pathways is frequently implicated in the pathophysiology of neurodevelopmental disorders.
Figure 2: Key signaling pathways studied in iPSC models of neurodevelopment. Pathways (yellow) govern critical functions (green), and their dysregulation is linked to disorders (red).
The synergistic advantages of patient-specific modeling, genetic background preservation, and unlimited expansion establish iPSC technology as a uniquely powerful platform for deconstructing the complex mechanisms underlying neurodevelopmental disorders. By providing a renewable source of patient-derived neural cells, this technology enables researchers to move beyond correlation to causation, linking genetic findings from GWAS to functional cellular and molecular phenotypes in a human context. As differentiation protocols become more sophisticated, yielding increasingly complex neural assemblies and organoids, and as scalable manufacturing processes improve, the fidelity and utility of these models will only increase. The continued application of iPSC-based models promises to accelerate the identification of novel therapeutic targets and the development of personalized interventions for individuals with neurodevelopmental conditions.
The study of neurodevelopmental disorders (NDDs) has been fundamentally transformed by the advent of induced pluripotent stem cell (iPSC) technology. This revolutionary approach enables researchers to generate patient-specific neural cells, providing unprecedented access to the cellular and molecular underpinnings of complex neurodevelopmental conditions. NDDs—including autism spectrum disorder (ASD), schizophrenia, and rare genetic syndromes—share common obstacles in research: limited access to functional human brain tissue, species-specific limitations of animal models, and profound genetic and clinical heterogeneity [17] [18]. iPSC technology effectively bypasses these constraints by allowing the reprogramming of patient somatic cells into pluripotent stem cells, which can subsequently be differentiated into disease-relevant neural cell types, including various neuronal subtypes, astrocytes, oligodendrocytes, and microglia [17] [3].
The application of iPSC models has been particularly valuable for disorders affecting the central nervous system, given the ethical limitations of working with human embryonic stem cells and the inability to sustainably maintain primary neural cultures [17]. Furthermore, iPSCs retain the complete genetic background of the donor, enabling researchers to investigate the complex interactions between genetic risk factors and environmental influences in NDD pathogenesis [17] [19]. The versatility of iPSC technology extends beyond simple two-dimensional (2D) monocultures to increasingly complex three-dimensional (3D) organoid systems that more faithfully recapitulate the architecture and cellular diversity of the developing human brain [17] [18]. This technical advancement has opened new avenues for elucidating disease mechanisms, identifying novel therapeutic targets, performing high-throughput drug screening, and developing personalized treatment approaches for NDDs [17] [3].
iPSC Modeling of Autism Spectrum Disorders
Autism spectrum disorder represents a group of complex neurodevelopmental conditions characterized by persistent deficits in social communication and interaction, alongside restricted, repetitive patterns of behavior, interests, or activities [17]. The etiopathogenesis of ASD is multifactorial, involving complex interactions between genetic and environmental factors, with heritability estimates ranging from 50% to 90% based on family and twin studies [17]. iPSC-based models have provided crucial insights into both syndromic (associated with known genetic conditions) and non-syndromic (idiopathic) forms of ASD by enabling the investigation of pathological mechanisms in disease-relevant human cell types [17] [19].
Key Cellular Phenotypes in ASD iPSC Models
Research using iPSC-derived neural progenitor cells (NPCs) from individuals with ASD and macrocephaly has revealed increased cellular proliferation resulting from alterations in a canonical Wnt-β-catenin/BRN transcriptional cascade [17]. These abnormalities in proliferation lead to aberrant neurogenesis and reduced synaptogenesis, ultimately contributing to functional defects in neuronal networks [17]. Similar hyperproliferation phenotypes have been observed in NPCs derived from individuals with syndromic forms of ASD, including Rett syndrome (RTT), Fragile X syndrome (FXS), tuberous sclerosis complex (TSC), Phelan-McDermid syndrome (PMDS), and Timothy syndrome (TS) [17].
In addition to proliferation defects, iPSC-derived neurons from ASD patients have demonstrated various synaptic abnormalities and electrophysiological alterations. These include impaired neurite outgrowth, aberrant synaptogenesis, and network hyperexcitability, which may underlie the behavioral manifestations observed in ASD [17] [19]. The ability to recapitulate these core cellular phenotypes in vitro has positioned iPSC technology as a powerful platform for identifying potential therapeutic interventions and conducting drug screening campaigns.
ASD Modeling Table
Table 1: Summary of Key Findings from iPSC-Based Studies of Autism Spectrum Disorders
| Disorder Category | Specific Disorder/Model | Key Cellular Phenotypes | Molecular Pathways Involved |
|---|---|---|---|
| Non-syndromic ASD | ASD with macrocephaly | NPC hyperproliferation, aberrant neurogenesis, reduced synaptogenesis | Wnt-β-catenin/BRN transcriptional cascade [17] |
| Syndromic ASD | Rett Syndrome (RTT) | NPC dysfunction, synaptic deficits, network abnormalities | MECP2 mutations, chromatin remodeling [17] |
| Fragile X Syndrome (FXS) | NPC hyperproliferation, aberrant neurite outgrowth, mGluR signaling defects | FMR1 silencing, mGluR pathway, AMPA receptor trafficking [17] [20] | |
| Tuberous Sclerosis Complex (TSC) | NPC hyperproliferation, neuronal hyperexcitability, dysregulated growth | mTOR signaling pathway [17] [18] | |
| Phelan-McDermid Syndrome (PMDS) | NPC dysfunction, synaptic deficits | SHANK3 mutations, synaptic scaffolding [17] | |
| Timothy Syndrome (TS) | NPC dysfunction, calcium signaling defects | CACNA1C mutations, calcium signaling [17] |
Elucidating Schizophrenia Mechanisms Through iPSC Models
While the search results provided limited specific information on schizophrenia modeling, the general principles of iPSC-based NDD research apply to this complex neuropsychiatric disorder. Schizophrenia is understood to have a significant neurodevelopmental component, with genetic and environmental factors interacting during critical periods of brain development to increase vulnerability to the disorder. iPSC models derived from patients with schizophrenia would likely focus on key processes such as neuronal migration, cortical patterning, synaptic formation and function, and the balance between excitatory and inhibitory neurotransmission.
The integration of iPSC technology with genomic approaches would be particularly valuable for schizophrenia research, given the polygenic nature of the disorder and the identification of numerous risk variants through genome-wide association studies. Isogenic lines generated using CRISPR/Cas9 technology could help determine the functional consequences of specific risk alleles in a controlled genetic background. Furthermore, the application of 3D brain organoid models could provide insights into potential cortical maldevelopment and disrupted cell positioning that may contribute to schizophrenia pathogenesis.
Rare Genetic Neurodevelopmental Syndromes
iPSC technology has proven particularly transformative for studying rare neurodevelopmental disorders, where the scarcity of patients, lack of neural tissues for analysis, and absence of representative animal models have historically hampered research progress [20]. These disorders, while individually rare, collectively affect a substantial proportion of the world's population and often present with severe neurological symptoms [20].
Fragile X Syndrome (FXS)
Fragile X syndrome represents the most common inherited form of intellectual disability and a frequent monogenic cause of ASD [20]. FXS results from an expansion of CGG repeats (>200 repeats) in the 5'-untranslated region of the FMR1 gene, leading to epigenetic silencing and loss of fragile X mental retardation protein (FMRP) [20]. iPSCs generated from FXS patients (FXS-iPSCs) maintain the hypermethylated, transcriptionally inactive state of the FMR1 gene, unlike embryonic stem cells where FMR1 downregulation occurs only during differentiation [20].
Neurons derived from FXS-iPSCs display aberrant differentiation characterized by defective neurite outgrowth, with impairments in both initiation and extension processes [20]. This suggests that normal FMRP expression is critical for early neurodevelopmental events preceding synaptogenesis. Additionally, NPCs derived from FXS-iPSCs show altered metabotropic glutamate receptor (mGluR) signaling, resulting in increased calcium influx and impaired differentiation [20]. Importantly, CRISPR/Cas9-mediated removal of CGG repeats or treatment with chromatin remodeling agents has demonstrated that the epigenetic silencing of FMR1 is reversible, with restoration of FMRP expression and rescue of neuronal differentiation abnormalities [20].
Rett Syndrome (RTT)
Rett syndrome is an X-linked neurodevelopmental disorder primarily affecting females, characterized by apparently normal early development followed by regression, loss of purposeful hand skills, gait abnormalities, and stereotypic hand movements [20]. RTT is predominantly caused by mutations in the MECP2 gene, which encodes methyl-CpG-binding protein 2, a critical regulator of gene expression and chromatin architecture. iPSC-derived neurons from RTT patients have revealed defects in neuronal maturation, synaptic density, and electrophysiological properties, providing insights into the consequences of MECP2 dysfunction in human neurons [17] [20].
Tuberous Sclerosis Complex (TSC)
Tuberous sclerosis complex is an autosomal dominant disorder characterized by benign tumors in multiple organs, including the brain, where it is associated with epilepsy, intellectual disability, and ASD features [18]. TSC results from mutations in either TSC1 or TSC2 genes, leading to constitutive activation of the mTOR signaling pathway. iPSC-derived neurons from TSC patients have demonstrated neuronal hyperexcitability and abnormal morphology, phenotypes that can be rescued by mTORC1-selective inhibitors [18]. These findings highlight the utility of iPSC models not only for understanding disease mechanisms but also for preclinical drug testing and therapeutic development.
Rare NDD Modeling Table
Table 2: Summary of iPSC-Based Studies of Rare Neurodevelopmental Disorders
| Rare NDD | Genetic Cause | Key iPSC-Derived Phenotypes | Therapeutic Insights |
|---|---|---|---|
| Fragile X Syndrome | FMR1 CGG repeat expansion (>200) | FMR1 hypermethylation, defective neurite outgrowth, altered mGluR signaling, increased calcium influx in NPCs [20] | CRISPR/Cas9-mediated CGG excision restores FMRP; chromatin remodeling agents reverse silencing [20] |
| Rett Syndrome | MECP2 mutations | NPC dysfunction, defects in neuronal maturation, reduced synaptic density, electrophysiological abnormalities [17] [20] | - |
| Tuberous Sclerosis Complex | TSC1 or TSC2 mutations | NPC hyperproliferation, neuronal hyperexcitability, abnormal morphology [17] [18] | mTORC1-selective inhibitors rescue hyperexcitability and morphological defects [18] |
| Phelan-McDermid Syndrome | SHANK3 mutations | NPC dysfunction, synaptic deficits | - |
| Timothy Syndrome | CACNA1C mutations | NPC dysfunction, calcium signaling defects | - |
Technical Methodologies in iPSC-Based NDD Research
iPSC Generation and Neural Differentiation
The fundamental process of generating iPSCs involves reprogramming somatic cells (typically fibroblasts or blood cells) through the expression of specific transcription factors. The original reprogramming factors—OCT4, SOX2, KLF4, and c-MYC (OSKM)—remain widely used, although alternative combinations such as OCT4, SOX2, NANOG, and LIN28 have also been employed successfully [3]. Following iPSC generation and characterization, several strategies exist for differentiating these pluripotent cells into neural lineages.
The two main approaches for generating neural cells from iPSCs include direct differentiation and reprogramming. In direct differentiation, iPSCs are first induced to form neural progenitor cells (NPCs) through dual SMAD inhibition, followed by exposure to specific trophic factors (e.g., cAMP, BDNF, NT3, and GDNF) to promote terminal differentiation into specific neuronal subtypes [17]. Alternatively, reprogramming strategies involve direct conversion using lentiviral delivery of neural-specific transcription factors coupled with antibiotic selection cassettes for efficient conversion and purification [17]. Each method presents distinct advantages and limitations, with the choice often depending on laboratory preference, desired neuronal subtype, and specific application requirements.
2D vs. 3D Modeling Approaches
Traditional iPSC-based disease modeling has relied primarily on two-dimensional (2D) monolayer cultures, which have proven valuable for investigating cell-autonomous phenotypes, high-content imaging, and electrophysiological characterization [17]. However, the recognition that human pathologies originate within the context of complex tissue environments has driven the development of three-dimensional (3D) brain organoid models [17] [18].
Brain organoids are self-organizing 3D aggregates derived from iPSCs that contain multiple neural cell types and develop cerebral-like structures, more closely mimicking the in vivo human brain microenvironment and pathophysiology [17]. These 3D models have been particularly informative for studying disorders involving cortical maldevelopment, disruptions in neuronal migration, and altered cell positioning, as they recapitulate important aspects of early developmental processes such as progenitor proliferation, neuronal migration, and layer formation [18]. The integration of 3D organoids with multi-omics approaches and advanced functional assays represents the cutting edge of iPSC-based NDD research.
Experimental Workflow Diagram
Diagram 1: Experimental Workflow for iPSC-Based NDD Research
Successful iPSC-based modeling of NDDs requires carefully selected reagents and methodologies. The table below outlines key resources essential for conducting robust iPSC studies in this field.
Table 3: Essential Research Reagents for iPSC-Based NDD Modeling
| Category | Specific Reagents/Tools | Function/Application | Technical Notes |
|---|---|---|---|
| Reprogramming Factors | OCT4, SOX2, KLF4, c-MYC (OSKM) or OCT4, SOX2, NANOG, LIN28 | Somatic cell reprogramming to pluripotency | Delivery via integrating (lentivirus) or non-integrating (episomal, mRNA) methods [3] |
| Neural Induction | Dual SMAD inhibitors (SB431542, LDN193189) | Efficient neural induction from iPSCs | Standard approach for NPC generation [17] |
| Neural Differentiation | BDNF, GDNF, NT3, cAMP | Terminal differentiation of NPCs into mature neurons | Concentration and timing vary by neuronal subtype [17] |
| Gene Editing | CRISPR/Cas9 systems, TALENs, ZFNs | Generation of isogenic controls; disease modeling of specific mutations | CRISPR/Cas9 most widely used for efficient genome engineering [20] |
| Characterization Antibodies | SSEA4, Tra-1-60, OCT3/4 (pluripotency); TUJ1, MAP2, NeuN (neuronal) | Validation of pluripotent state and neural differentiation | Essential for quality control throughout differentiation process [21] |
| Functional Assays | Multi-electrode arrays, calcium imaging, patch clamp | Electrophysiological characterization of neuronal function | Critical for assessing functional phenotypes in NDD models [18] |
Signaling Pathways in Neurodevelopmental Disorders
iPSC-based studies have identified several key signaling pathways that are commonly disrupted across multiple NDDs. The diagram below illustrates some of these critical pathways and their interconnections.
Diagram 2: Key Signaling Pathways in Neurodevelopmental Disorders
iPSC technology has fundamentally transformed our approach to studying neurodevelopmental disorders, providing unprecedented access to patient-specific neural cells and enabling mechanistic insights that were previously inaccessible. The applications spanning autism spectrum disorder, schizophrenia, and rare genetic syndromes demonstrate the remarkable versatility of this platform for both basic research and therapeutic development. The integration of iPSC-derived models with advanced functional assays, multi-omics approaches, and computational analytics represents the current frontier in NDD research [18].
Despite substantial progress, challenges remain in standardizing differentiation protocols, improving reproducibility across laboratories, and effectively integrating the massive multi-layered datasets generated by these sophisticated models [18]. Furthermore, translating findings from cellular models into clinical interventions will require close collaboration between basic scientists, clinicians, and computational experts. Nevertheless, the trajectory is clear: by uniting stem cell biology, multi-omics integration, and computational frameworks, the field is moving toward more predictive, patient-specific, and ultimately actionable models of neurodevelopmental disorders. The continued refinement of iPSC-based technologies holds exceptional promise for unraveling the complex etiology of NDDs and developing effective targeted therapies for these debilitating conditions.
Advanced iPSC Methodologies: From 2D Cultures to 4D Multi-Organ Systems for Neural Research
The study of human neurodevelopmental disorders has long been constrained by the limited availability of authentic human tissue models that accurately recapitulate the complexity of the developing brain. Traditional two-dimensional (2D) monolayer cultures, while valuable for specific applications, fail to capture the three-dimensional architecture and cell-cell interactions inherent to native neural tissue. The advent of induced pluripotent stem cell (iPSC) technology has revolutionized this landscape, enabling the derivation of patient-specific neural cells and the development of increasingly sophisticated three-dimensional (3D) model systems [3]. These advances have been particularly transformative for researching neurodevelopmental disorder mechanisms, as they provide unprecedented access to human-specific developmental processes and disease phenotypes that are often not faithfully reproduced in animal models [18].
The progression from 2D monolayers to 3D organoids and recently to assembloids represents a paradigm shift in how researchers model the intricate cellular relationships and spatial organization critical to understanding brain development and dysfunction. Organoids are defined as 3D structures derived from stem cells that self-organize through cell sorting and spatially restricted lineage commitment, recapitulating aspects of the developing organ [22]. Assembloids represent a further advancement, created by facing regionally specified organoids to model interactions between different brain areas or cell types [4]. This evolution in model systems has created new opportunities to dissect the cellular and molecular mechanisms underlying neurodevelopmental disorders, leveraging the genetic background of patients while overcoming the limitations of earlier modeling approaches.
Technical Foundations and Methodological Approaches
iPSC Generation and Neural Differentiation
The foundation of all modern human neural tissue models begins with the generation of induced pluripotent stem cells (iPSCs). Since the groundbreaking work of Takahashi and Yamanaka in 2006, iPSC technology has matured significantly, with numerous refinements in reprogramming methods, factor delivery, and somatic cell source selection [3] [23]. Current approaches prioritize non-integrating delivery methods such as Sendai virus, episomal plasmids, or mRNA transfection to minimize genomic alteration risks while maintaining high reprogramming efficiency [23]. The selection of somatic cell sources has expanded beyond the original dermal fibroblasts to include more accessible options such as peripheral blood mononuclear cells (PBMCs) and renal epithelial cells from urine, facilitating the creation of patient-specific models from diverse clinical populations [23].
The differentiation of iPSCs into neural lineages typically employs dual SMAD inhibition to direct cells toward a neural fate, followed by region-specific patterning factors that generate distinct neuronal subtypes [24]. For cortical excitatory neurons, this involves activation of Wnt and FGF signaling, while ventral forebrain identities (including interneurons) require SHH activation. The efficiency and reproducibility of these differentiation protocols have improved substantially, enabling the generation of highly enriched populations of specific neuronal subtypes, though heterogeneity remains a challenge, particularly in more complex 3D systems [23].
2D Monolayer Culture Systems
Two-dimensional monolayer cultures represent the most reductionist approach to modeling neural tissues in vitro. In these systems, iPSCs are differentiated into neural progenitor cells (NPCs) and subsequently into neurons or glia on flat, coated surfaces, typically in the presence of specific patterning factors. The methodology involves several sequential steps: initial neural induction via dual SMAD inhibition, regional patterning using small molecules or growth factors, terminal differentiation through neurotrophic factor withdrawal, and finally functional maturation over several weeks [23].
The key advantage of 2D systems lies in their simplicity, reproducibility, and scalability, making them ideal for high-throughput screening applications and reductionist experimental designs. However, this simplicity comes with significant limitations, as 2D cultures lack the complex cytoarchitecture, cell-cell interactions, and microenvironmental cues present in the developing brain [25] [22]. Studies comparing transcriptomic profiles between 2D monolayers and 3D organoids have revealed profound differences in gene expression patterns, with monolayers exhibiting suppressed Notch signaling and altered radial glia polarity that ultimately impairs the generation of intermediate progenitors and cortical neurons [25].
3D Organoid Culture Systems
Three-dimensional organoid cultures represent a significant advancement in neural tissue modeling by recapitulating aspects of the brain's spatial organization and developmental processes. Cerebral organoids can be generated through multiple approaches, including unguided methods that rely on intrinsic self-organization capacity and guided protocols that use exogenous patterning factors to generate region-specific organoids [26]. The basic protocol involves forming embryoid bodies from iPSCs, inducing neural differentiation, embedding these structures in extracellular matrix (typically Matrigel), and maintaining them in spinning bioreactors or orbital shakers to enhance nutrient and oxygen exchange [26] [22].
Organoid cultures model several key aspects of human brain development, including the emergence of ventricular zones, the generation of diverse neuronal subtypes, and the formation of layered cortical structures. The self-organization capacity of organoids arises from cell sorting and spatially restricted lineage commitment, processes that mimic in vivo development [22]. However, organoids face challenges including necrotic core formation due to limited nutrient diffusion, heterogeneity in size and cellular composition, and incomplete representation of later developmental stages [26]. Recent advances have addressed some limitations through the incorporation of microfluidic systems to improve vascularization, enhanced patterning protocols for more reproducible regional specification, and extended culture durations to capture later developmental events [4] [26].
Assembloid Culture Systems
Assembloids represent the most advanced in vitro model for neural tissues, created by fusing regionally specified organoids to study interactions between different brain areas or cell types. This approach enables researchers to model long-distance migration, circuit formation, and interactions between distinct neuronal populations that cannot be captured in single organoids [4]. A representative protocol involves generating dorsal (cortical) and ventral (ganglionic eminence) organoids separately using specific patterning factors, then bringing them together at a defined developmental stage to form assembloids, with subsequent analysis of interneuron migration and integration [4].
The assembloid platform has enabled groundbreaking studies of human-specific developmental processes, such as the prolonged migration of interneurons that continues into the postnatal period in humans. Recent work has demonstrated that after extended culture (over 200 days), newly born migratory interneurons in assembloids arrange themselves into connected chains surrounded by astrocytes, essentially recapitulating the architectural and migratory patterns observed in early postnatal human brains [4]. This model system provides unprecedented opportunities to study cellular interactions in a human-specific context, particularly for neurodevelopmental disorders involving aberrant migration or circuit formation.
Comparative Analysis of Model Systems
Architectural and Cellular Complexity
The architectural fidelity of neural tissue models profoundly influences their cellular composition and developmental potential. Comparative studies between 2D monolayers and 3D organoids have revealed striking differences in their cellular organization and differentiation capacity.
Table 1: Architectural and Cellular Features of Neural Tissue Models
| Feature | 2D Monolayers | 3D Organoids | Assembloids |
|---|---|---|---|
| Spatial Organization | Flat, uniform structure | Self-organized 3D structures with regional zones | Multiple integrated regions with interface zones |
| Cell-Cell Interactions | Limited to horizontal plane | Multi-directional, including apical-basal polarity | Cross-regional interactions and long-distance migration |
| Radial Glia Polarity | Altered, disrupted polarity | Preserved apical-basal polarity with ventricular zones | Preserved polarity across multiple regions |
| Neuronal Differentiation | Impaired generation of intermediate progenitors and cortical neurons | Sequential generation of deep and upper layer neurons | Region-specific neuronal subtypes with functional connectivity |
| Glial Populations | Limited astrocyte and microglia incorporation | Astrocytes present, microglia often missing | Multiple glial types supporting neuronal migration and function |
| Vascularization | Absent | Absent, leading to necrotic cores | Limited, but improved nutrient exchange at interfaces |
Research by Scuderi et al. directly comparing 2D monolayers and 3D organoids demonstrated that organoids exhibit more efficient Notch signaling in ventricular radial glia due to preserved cell adhesion, resulting in subsequent generation of intermediate progenitors and outer radial glia in a sequence that better recapitulates cortical development [25]. Network analyses revealed co-clustering of cell adhesion and Notch-related transcripts in a module strongly downregulated in monolayers, providing a molecular explanation for their limited differentiation capacity [25].
Assembloids further enhance this architectural complexity by modeling interactions between different brain regions. For example, dorsal-ventral assembloids have revealed that chain migration of interneurons requires both intrinsic cues from late-born interneurons and specific interactions with surrounding astrocytes [4]. This level of cellular crosstalk cannot be modeled in simpler systems and provides critical insights into human-specific developmental processes that may be disrupted in neurodevelopmental disorders.
Physiological and Functional Properties
The functional properties of neurons and neural circuits differ significantly across model systems, with important implications for their utility in disease modeling and drug screening.
Table 2: Functional Properties of Neural Tissue Models
| Property | 2D Monolayers | 3D Organoids | Assembloids |
|---|---|---|---|
| Neural Activity | Synchronized bursting; limited network complexity | Spontaneous, synchronized network activity with complex bursting patterns | Cross-regional synchronized activity; developing circuit dynamics |
| Synaptic Development | Immature synapses; limited pruning | Maturing synapses with spontaneous pruning | Region-specific synaptic properties; functional connectivity between regions |
| Circuit Formation | Limited local connectivity | Local microcircuits with layered organization | Long-range connections between different regions |
| Metabolic Characteristics | Uniform nutrient and oxygen access | Gradient-dependent metabolism with hypoxic cores | Improved metabolic support at interfaces |
| Response to Stimulation | Homogeneous response to pharmacological agents | Stratified responses based on spatial position | Region-specific responses to modulators |
| Disease Modeling Fidelity | Limited to cell-autonomous phenotypes | Captures some tissue-level pathologies | Models non-cell-autonomous mechanisms and circuit-level defects |
3D culture systems more closely resemble the architectural and functional properties of in vivo tissues, with cells exposed to different concentrations of nutrients, growth factors, and oxygen depending on their localization [22]. This spatial heterogeneity creates microenvironments that influence cellular phenotypes, including differential responses to pharmacological agents that may explain discrepancies between traditional drug screening results and clinical outcomes [22]. For instance, studies have demonstrated that temozolomide resistance in glioblastoma 3D cultures was 50% higher than in 2D models, highlighting the importance of microenvironmental context for therapeutic response [22].
Functional analyses of 3D neurospheres have revealed reliable spontaneous activity that offers functional tissue culture readouts of neural firing, including oscillatory network activity that becomes more complex with maturation [24]. Assembloids exhibit even more sophisticated functional properties, with emerging evidence of coordinated activity between fused regions that begins to approximate developing neural circuits in the human brain [4].
Experimental Utility and Limitations
Each model system offers distinct advantages and limitations for specific research applications in neurodevelopmental disorders.
Table 3: Experimental Applications of Neural Tissue Models
| Application | 2D Monolayers | 3D Organoids | Assembloids |
|---|---|---|---|
| High-Throughput Screening | Excellent for scalability and reproducibility | Moderate throughput with emerging platforms | Low throughput, technically challenging |
| Genetic Manipulation | Highly efficient via viral transduction or CRISPR | Moderate efficiency with improved techniques | Limited to pre-fusion manipulation or viral delivery |
| Live Imaging | Straightforward with full optical access | Challenging due to opacity and depth | Complex, requiring specialized microscopy |
| Transcriptomic Analysis | Homogeneous cell populations | Heterogeneous populations requiring spatial methods | Extreme heterogeneity with regional identities |
| Disease Modeling Scope | Cell-autonomous mechanisms | Tissue-level phenotypes and cell non-autonomous effects | Circuit-level disorders and long-range interactions |
| Reproducibility | High consistency across experiments | Moderate, with batch-to-batch variability | Lower, with fusion efficiency variations |
| Developmental Timeframe | Limited to early maturation stages | Extended development, but incomplete maturation | Longest culture duration with advanced maturation |
For genetic studies and high-throughput drug screening, 2D monolayers remain the system of choice due to their scalability and reproducibility. The "village editing" approach exemplifies this utility, where CRISPR/Cas9 gene editing in a cell village format enabled efficient NRXN1 knockout across iPSC lines from 15 donors with varying polygenic risk scores for schizophrenia [4]. Such large-scale, genetically diverse studies would be prohibitively challenging in more complex 3D systems.
However, for modeling non-cell-autonomous disease mechanisms and circuit-level defects, assembloids provide unique capabilities. The extended culture durations possible with assembloids (up to 390 days in some reports) enable modeling of postnatal developmental events that were previously inaccessible in vitro [4]. This extended timeline is particularly valuable for neurodevelopmental disorders with postnatal onset or progression, allowing researchers to capture developmental processes that occur over extended periods in the human brain.
Signaling Pathways in Neural Development Models
The signaling pathways active in neural tissue models significantly influence their developmental trajectory and cellular composition. Comparative studies have revealed fundamental differences in how these pathways are engaged across different culture systems.
Diagram 1: Signaling Pathways in 2D vs 3D Neural Models
The diagram illustrates key signaling differences between 2D and 3D neural models. In 3D organoids, preserved cell-cell adhesion enables robust Notch signaling, maintaining radial glia in a progenitor state and supporting sequential generation of neuronal subtypes [25]. In contrast, 2D monolayers exhibit suppressed Notch signaling due to disrupted cell adhesion, leading to precocious differentiation and impaired generation of intermediate progenitors and outer radial glia [25]. Integrin signaling is enhanced in 2D systems, promoting increased proliferation but altering the normal developmental sequence of neurogenesis.
These signaling differences have profound implications for modeling neurodevelopmental disorders. For example, the village editing study of NRXN1 deletions in multiple genetic backgrounds found that genetic background deeply influences gene expression changes in NRXN1 knockout neurons, highlighting the importance of capturing gene-gene interactions in neurodevelopmental disorder models [4]. The more physiologically relevant signaling environment in 3D systems may provide a more accurate context for evaluating such genetic interactions and their contribution to disease risk.
Applications in Neurodevelopmental Disorder Research
Disease Modeling with Patient-Specific iPSCs
The integration of iPSC technology with neural tissue models has created unprecedented opportunities for studying neurodevelopmental disorders with patient-specific genetic backgrounds. This approach allows researchers to retain the complete genetic architecture of patients, providing direct access to disease mechanisms in a human context [18]. Studies of Tuberous Sclerosis Complex (TSC) have leveraged hiPSC-derived neurons to interrogate dysregulation of the mTOR signaling pathway, leading to the development of mTORC1-selective inhibitors that rescue hyperexcitability and abnormal neuronal morphology in TSC2−/− neurons [18]. Similarly, research on hereditary sensory and autonomic neuropathy type IV (HSAN IV) using patient-derived dorsal root ganglion (DRG) organoids revealed that NTRK1 mutations disrupt the balance of neuronal and glial differentiation in human DRG during development, providing mechanistic insights that could not be obtained from animal models [4].
The capacity to correct disease-causing mutations using CRISPR-based gene editing further enhances the utility of these models by enabling isogenic control generation. In the HSAN IV study, researchers generated an isogenic control by correcting the patient's mutation, confirming that observed phenotypes were directly attributable to the NTRK1 mutation rather than background genetic variation [4]. This rigorous experimental design strengthens conclusions about disease mechanisms and provides a robust platform for therapeutic development.
High-Content Screening and Therapeutic Development
Advanced neural tissue models are increasingly being applied in drug discovery and development pipelines, offering more physiologically relevant systems for evaluating candidate therapeutics. The integration of these models with high-content imaging, multi-electrode array recording, and transcriptomic profiling enables comprehensive assessment of drug effects on complex cellular systems [18].
For neurodevelopmental disorders involving synaptic dysfunction, such as schizophrenia and autism spectrum disorders, the enhanced synaptic development and network activity in 3D systems provides a more relevant context for evaluating potential therapeutics. The village editing approach for studying NRXN1 deletions exemplifies how complex genetic interactions can be modeled at scale, enabling researchers to assess how genetic background influences response to perturbations [4]. This capability is particularly valuable for neurodevelopmental disorders with complex genetic architecture, where therapeutic response may depend on an individual's unique combination of risk variants.
The FDA Modernization Act 2.0, which permits cell-based assays as an alternative to animal testing for drug and biological product applications, has further accelerated the adoption of iPSC-derived models in pharmaceutical development [27]. Companies like bit.bio now offer researchers the ability to order specific mutations in opti-ox powered human-iPSC derived cells, providing consistent, defined, cryopreserved products that can be incorporated into drug discovery workflows [27]. These advances are making human-specific models increasingly accessible and standardized, facilitating their integration into therapeutic development pipelines.
Research Reagent Solutions
The successful implementation of neural tissue models requires specialized reagents and materials tailored to the unique requirements of each system. The table below summarizes key research reagents and their applications across different model types.
Table 4: Essential Research Reagents for Neural Tissue Models
| Reagent Category | Specific Examples | Function | Application Across Models |
|---|---|---|---|
| Reprogramming Factors | OCT3/4, SOX2, KLF4, c-MYC (OSKM); OCT3/4, SOX2, NANOG, LIN28 | Induction of pluripotency in somatic cells | Foundation for all iPSC-derived models |
| Small Molecule Inhibitors | CHIR99021 (GSK3β inhibitor), PD0325901 (MEK inhibitor), A-83-01 (TGF-β receptor inhibitor), Thiazovivin (ROCK inhibitor) | Direct differentiation toward neural lineages; enhance reprogramming efficiency | Used in 2D differentiation and 3D patterning |
| Extracellular Matrix | Matrigel, laminin, collagen-based hydrogels | Provide 3D scaffold for structural support and signaling cues | Essential for organoid and assembloid culture |
| Growth Factors | EGF, FGF, noggin (BMP inhibitor), R-spondin-1 (Wnt agonist), BDNF, GDNF | Regional patterning and support of neuronal survival/maturation | Concentration and timing critical for regional specification |
| Neural Differentiation Supplements | B27, N2, N21, neurobasal media | Provide essential nutrients and signaling molecules for neural cells | Standard component across all neural culture systems |
| Gene Editing Tools | CRISPR/Cas9 systems, piggyBac transposons, episomal plasmids | Introduction of disease-associated mutations or correction of existing mutations | Enables isogenic control generation and disease modeling |
| Functional Assay Reagents | GCaMP6f (calcium indicator), adeno-associated viruses (AAV), multielectrode arrays | Monitoring neural activity and network dynamics | Critical for functional characterization across models |
The selection of appropriate reagents is critical for model fidelity and reproducibility. For example, the choice of extracellular matrix significantly influences organoid development, with Matrigel remaining the most common choice despite batch-to-batch variability [22]. Emerging synthetic matrices offer improved consistency and tunability, potentially addressing one of the key sources of variability in organoid research. Similarly, the development of defined neural differentiation supplements like B27 and N2 has greatly enhanced the reproducibility of neural differentiation across laboratories, though differences in formulation can still influence results [24] [23].
Future Perspectives and Concluding Remarks
The evolution from 2D monolayers to 3D organoids and assembloids represents a transformative progression in how researchers model human neural development and its disorders. Each model system offers unique advantages and limitations, making them complementary rather than mutually exclusive tools for investigating different aspects of neurodevelopmental processes. 2D monolayers remain invaluable for high-throughput screening and reductionist experimental designs, while 3D organoids provide more physiologically relevant context for studying tissue-level phenotypes, and assembloids enable investigation of circuit-level interactions between different brain regions.
Despite remarkable progress, significant challenges remain in the development and application of these models. Standardization of differentiation protocols, improvement of reproducibility across laboratories, and enhancement of maturation states continue to be pressing issues [18]. The integration of vascular components, immune cells, and sensory interfaces represents the next frontier in model complexity, potentially enabling even more faithful recapitulation of the in vivo environment [26]. Additionally, the development of analytical methods capable of resolving the extreme heterogeneity of these systems, particularly assembloids, requires continued innovation in spatial transcriptomics, live imaging, and computational analysis.
The application of these advanced models to neurodevelopmental disorder research holds tremendous promise for elucidating disease mechanisms and developing novel therapeutics. By combining patient-specific iPSCs with increasingly sophisticated tissue engineering approaches, researchers can now model complex genetic interactions, non-cell-autonomous disease mechanisms, and circuit-level dysfunction in a human-specific context. As these technologies continue to evolve and become more accessible, they are poised to fundamentally advance our understanding of neurodevelopmental disorders and accelerate the development of effective interventions.
In the study of neurodevelopmental disorders (NDDs), human induced pluripotent stem cell (iPSC) models have emerged as a transformative platform, enabling direct investigation of patient-derived neurons and glia. A central challenge in this field, however, is distinguishing disease-causing pathological mechanisms from the confounding effects of inherent genetic background variability. Isogenic controls—genetically identical iPSC lines that differ only at a specific disease-relevant locus—represent the methodological gold standard for addressing this challenge. The emergence of CRISPR/Cas9 genome editing has dramatically accelerated the creation of these precision models, allowing researchers to introduce or correct disease-associated mutations with unprecedented efficiency in diverse genetic backgrounds.
This technical guide examines contemporary protocols and considerations for employing CRISPR/Cas9 in iPSCs to generate isogenic controls for NDD research. We detail an optimized high-efficiency editing method, analyze the landscape of editing outcomes with a focus on safety and validation, and present applications in advanced neural model systems such as brain organoids and assembloids. The integration of these genome engineering approaches with complex iPSC-derived models is pushing forward our capacity to decipher the molecular and cellular underpinnings of disorders including schizophrenia, autism spectrum disorder, and hereditary sensory neuropathies within a genetically controlled framework.
Optimized Workflow for High-Efficiency CRISPR Editing in iPSCs
Conventional CRISPR/Cas9 editing in iPSCs often faces limitations such as low homologous recombination efficiency, significant cell death following single-cell cloning, and the extensive time investment required to isolate and validate clonal lines. Recent methodological advancements have systematically addressed these bottlenecks by targeting key cellular stress and repair pathways.
Core Protocol: Enhanced Homology-Directed Repair (HDR)
An optimized protocol demonstrates that combining p53 pathway inhibition with pro-survival small molecules can achieve homologous recombination rates exceeding 90% in human iPSCs, a substantial improvement over base protocol efficiencies of approximately 3% [28].
The mechanistic rationale for this multi-factor approach is twofold:
- Minimizing CRISPR-Induced Apoptosis: The Cas9 nuclease induces double-strand breaks (DSBs) that trigger p53-mediated apoptosis [28] [29]. Transient inhibition of this pathway, achieved here via transfection with a plasmid encoding shRNA against p53 (pCXLE-hOCT3/4-shp53-F), allows edited cells to survive the DNA damage response.
- Enhancing Cell Survivability: The physical stress of nucleofection and the subsequent single-cell cloning stage incur massive cell death. Supplementing culture media with a cocktail of pro-survival factors, including a ROCK inhibitor (Revitacell) and the cloning supplement CloneR, significantly improves the survival and recovery of edited cells [28].
Table 1: Key Reagents for High-Efficiency iPSC Genome Editing
| Reagent Category | Specific Reagent | Function & Rationale |
|---|---|---|
| Nuclease System | Alt-R S.p. HiFi Cas9 Nuclease V3 [28] | High-fidelity Cas9 variant; reduces off-target effects while maintaining robust on-target activity. |
| Guide RNA | Synthetic sgRNA (IDT) [28] | Defines genomic target sequence; complexed with Cas9 to form Ribonucleoprotein (RNP) for editing. |
| Repair Template | Single-Stranded Oligonucleotide (ssODN) [28] | Contains the desired point mutation and homologous arms for HDR; often includes a silent "PAM-disrupting" mutation to prevent re-cleavage. |
| p53 Inhibition | pCXLE-hOCT3/4-shp53-F plasmid [28] | shRNA-mediated transient p53 knockdown; prevents Cas9-induced apoptosis and enriches for HDR-edited cells. |
| Pro-Survival Supplements | CloneR, Revitacell [28] | Small molecule cocktails that inhibit apoptosis and enhance single-cell survival post-nucleofection and during cloning. |
| HDR Enhancer | Commercial HDR Enhancer (e.g., from IDT) [28] | Chemical additive that modestly shifts DNA repair balance towards HDR and away from error-prone NHEJ. |
Quantitative Outcomes of the Optimized Protocol
The efficacy of this optimized protocol is demonstrated by its application across multiple independent genetic loci and iPSC lines, showing consistent and dramatic improvements in HDR efficiency [28].
Table 2: Editing Efficiency Across Genetic Loci and Cell Lines
| Target Gene / Mutation | Base Protocol HDR Efficiency | Optimized Protocol HDR Efficiency | Fold Improvement |
|---|---|---|---|
| EIF2AK3 (rs867529) | 2.8% | 59.5% | 21x |
| EIF2AK3 (rs13045) | 4% | 25% | 6x |
| APOE R136S (PS 1.9.1 line) | Not Reported | 49% (Bulk); 100% (Clones) | - |
| PSEN1 E280A (Correction) | Not Reported | 97-98% (Bulk); 100% (Clones) | - |
This workflow significantly reduces the time and resources required to generate and isolate isogenic clones, compressing the timeline to as little as 8 weeks from nucleofection to validated clonal line [28].
Diagram 1: High-efficiency CRISPR workflow for isogenic iPSC line generation.
Beyond Efficiency: Assessing Genomic Integrity and Safety
While achieving high editing efficiency is a primary goal, a comprehensive safety assessment of the resulting clonal lines is equally critical, particularly for downstream disease modeling and therapeutic applications. Beyond routine checks for off-target mutations, recent studies highlight the pressing concern of on-target structural variations (SVs) induced by CRISPR/Cas9 editing [29].
The Spectrum of Unintended On-Target Edits
The genotoxic potential of CRISPR-induced double-strand breaks extends far beyond small insertions or deletions (indels). Deep genomic analyses have revealed that large-scale deletions (kilobase- to megabase-scale), chromosomal truncations, and even chromosomal translocations can occur at the target site with notable frequency [29]. These SVs are particularly concerning because they can delete critical regulatory elements or entire genes adjacent to the intended target, potentially confounding experimental results or posing oncogenic risks in a therapeutic context.
A critical technical note is that these large deletions are often "invisible" to standard PCR-based validation methods (e.g., Sanger sequencing or short-read amplicon sequencing). If a deletion removes one or both primer binding sites, the allele will fail to amplify, leading to an overestimation of HDR efficiency and an underestimation of aberrant editing outcomes [29].
Mitigation Strategies for Safer Editing
- Avoid DNA-PKcs Inhibition: The use of DNA-PKcs inhibitors (e.g., AZD7648) to enhance HDR rates has been shown to drastically increase the frequency of megabase-scale deletions and chromosomal translocations, by a factor of up to 1000-fold in some cases [29]. Their use in iPSC editing is not recommended.
- Leverage Advanced Analysis Tools: Employ SV detection methods such as CAST-Seq or LAM-HTGTS during the validation phase of candidate clones to identify those with clean on-target edits [29].
- Prioritize High-Fidelity Nucleases: Using high-fidelity Cas9 variants (e.g., HiFi Cas9) reduces off-target activity but does not eliminate on-target SVs. The optimized protocol described in Section 2.1 utilizes HiFi Cas9 V3 to this end [28].
- Conduct Comprehensive Clone Validation: A robust validation pipeline for a candidate isogenic clone should include: Karyotyping (G-banding) to check for gross chromosomal abnormalities [28]; Whole Genome Sequencing (WGS) to detect SVs and potential off-target sites [28]; and functional assays to confirm the expected phenotypic changes.
Diagram 2: CRISPR DNA repair pathways and structural variation risks.
Applications in Neurodevelopmental Disorder Research
The power of CRISPR-engineered isogenic iPSCs is fully realized when these models are differentiated into complex, physiologically relevant neural systems. This approach allows for the precise dissection of disease mechanisms in a human neural context.
Modeling Schizophrenia Genetic Risk in a Multi-Donor Framework
The study of schizophrenia (SCZ), a highly polygenic disorder, requires methodologies that can account for the influence of genetic background. A novel "village editing" approach was developed to address this [4]. Researchers performed CRISPR/Cas9-mediated knockout of the SCZ-associated gene NRXN1 in a "village" of iPSCs from 15 different donors, each with varying polygenic risk scores for SCZ. This powerful design enabled the simultaneous generation of heterozygous (33.1%) and homozygous (28.4%) NRXN1 deletions across most donor lines. Subsequent differentiation into cortical excitatory neurons and transcriptomic analysis revealed that the genetic background profoundly influenced the molecular consequences of NRXN1 loss. This establishes a new framework for studying the interaction of major effect risk genes with polygenic background in complex psychiatric disorders [4].
Elucidating Disease Mechanisms in 3D Organoid and Assembloid Models
The integration of isogenic iPSCs with 3D brain organoid technologies provides an unparalleled platform for studying NDDs in a tissue-like environment that recapitulates key aspects of human brain development [30].
Hereditary Sensory and Autonomic Neuropathy Type IV (HSAN IV): Researchers established a human dorsal root ganglion (DRG) organoid model of HSAN IV using patient-derived iPSCs carrying mutations in the NTRK1 gene [4]. They then used CRISPR to generate an isogenic control line by correcting the pathogenic mutations. Comparative analysis revealed that the patient organoids underwent a faulty lineage switch, with a marked reduction in sensory neurons and premature initiation of gliogenesis. This phenotype was rescued in the corrected isogenic line, directly linking the NTRK1 mutation to a disruption in the neuronal-glial differentiation balance during human sensory neuron development [4].
Studying Interneuron Migration in Assembled Tissues: Cortical assembloids, created by fusing region-specific organoids (e.g., dorsal and ventral forebrain organoids), can model the migration of interneurons observed in the postnatal human brain [4] [30]. These intricate models, which can be cultured for over a year, have revealed that this unique migratory process requires intrinsic cues from late-born interneurons and specific interactions with surrounding astrocytes [4]. This system provides a genetically controllable model to investigate how NDD-associated mutations disrupt the formation of complex neural circuits.
Table 3: iPSC-Derived Neural Models for NDD Research
| Model System | Key Features | Applications in NDD Research | Examples |
|---|---|---|---|
| 2D Neuronal Cultures | Monolayer cultures of specific neuronal subtypes; relatively simple and high-throughput. | Electrophysiology, high-content drug screening, synaptic studies. | Cortical excitatory neurons [4] |
| Brain Organoids | 3D, self-organizing structures that mimic the cellular diversity and architecture of developing brain regions. | Modeling neurogenesis, cell fate decisions, and the effect of mutations on tissue-level organization. | DRG organoids [4], Whole-brain & region-specific organoids [30] |
| Assembloids | Fusion of organoids from different brain regions to model inter-regional connectivity. | Studying cell migration, axon pathfinding, and neural circuit formation and dysfunction. | Cortical-striatal assembloids [30] |
The Scientist's Toolkit: Essential Reagents and Solutions
Successful genome engineering in iPSCs requires a suite of reliable, high-quality reagents. The following table catalogs essential solutions utilized in the protocols and applications discussed in this guide.
Table 4: Research Reagent Solutions for CRISPR/iPSC Workflows
| Reagent / Solution | Supplier Examples | Critical Function in Workflow |
|---|---|---|
| Alt-R HDR Enhancer | Integrated DNA Technologies (IDT) [31] | Boosts HDR efficiency up to 2-fold in hard-to-edit cells like iPSCs and HSPCs, without raising off-target edits. |
| Alt-R S.p. HiFi Cas9 | Integrated DNA Technologies (IDT) [28] | Engineered Cas9 nuclease with reduced off-target effects while maintaining high on-target activity. |
| Lipid Nanoparticles (LNPs) | Acuitas Therapeutics [32] [33] | Delivery vehicle for in vivo CRISPR therapy; has affinity for liver cells and allows for re-dosing. |
| ReproCardio (iPSC-Cardiomyocytes) | REPROCELL [27] | Commercially available human iPSC-derived cardiomyocytes for disease modeling and cardiotoxicity testing. |
| FeBO Culture System | N/A [30] | Protocol for generating brain organoids directly from fetal brain tissue, preserving native cellular diversity. |
| Matrigel | Corning [28] [30] | Basement membrane extract used as a substrate for feeder-free iPSC culture and for embedding organoids. |
| StemFlex/mTeSR Plus Media | Gibco/STEMCELL Technologies [28] | Chemically defined, xeno-free media for the maintenance and expansion of human iPSCs under feeder-free conditions. |
Functional phenotyping represents a critical approach in neuroscience for quantifying the dynamic activity of neurons and neural networks. In the context of researching neurodevelopmental disorders (NDDs) using induced pluripotent stem cell (iPSC) models, it provides direct insight into how genetic and molecular abnormalities translate into dysfunctional cellular signaling and communication. The integration of patient-derived iPSCs with advanced functional phenotyping techniques has opened new avenues for understanding the pathophysiology of rare NDDs, overcoming previous limitations posed by the scarcity of neural tissues and the inadequate recapitulation of human-specific processes in animal models [20] [18]. This technical guide details three cornerstone methodologies—patch clamp electrophysiology, multi-electrode array (MEA) recording, and calcium imaging—for the functional analysis of iPSC-derived neuronal models. It provides standardized protocols, quantitative comparisons, and practical frameworks for their application in deciphering the mechanistic underpinnings of NDDs.
The following table summarizes the core attributes, applications, and outputs of the three primary functional phenotyping techniques.
Table 1: Core Functional Phenotyping Techniques for iPSC-Derived Neuronal Models
| Technique | Primary Measurement | Spatial Resolution | Temporal Resolution | Key Applications in NDD Research | Primary Outputs |
|---|---|---|---|---|---|
| Patch Clamp Electrophysiology | Voltage-gated or ligand-gated currents across an individual neuron's membrane [34]. | Single-cell (micrometer scale) | Very High (microsecond scale) [35] | Characterization of cell-type-specific synaptic and cellular parameters; analysis of action potential properties, synaptic transmission, and ion channel function [34] [36]. | Action potential waveforms, postsynaptic currents (PSCs), membrane capacitance, input resistance. |
| Multi-Electrode Array (MEA) | Extracellular field potentials from a population of cells [35]. | Multi-cell / Network (tens to hundreds of micrometers) | High (millisecond scale) [35] | Long-term, non-invasive monitoring of network development and activity; assessment of synchronous bursting, spike rates, and oscillatory rhythms; high-throughput drug screening [34] [35]. | Spike raster plots, mean firing rate, burst characteristics, network synchrony indices. |
| Calcium Imaging | Fluctuations in intracellular calcium concentration ((Ca^{2+})) as a proxy for neuronal activation [37]. | Single-cell to network level (micrometer scale) | Moderate (millisecond to second scale) | Functional characterization of neuronal phenotypes; mapping of network connectivity; analysis of signaling amplitudes, duration, and oscillation frequency [37] [38]. | Fluorescence traces ((\Delta F/F)), calcium transient kinetics, spike inference, functional connectivity maps. |
Detailed Methodologies and Protocols
Patch Clamp Electrophysiology in Acute Brain Slices and iPSC-Derived Cultures
Patch clamp electrophysiology is considered the gold standard for detailed biophysical characterization of individual neurons. While historically used in acute brain slices, it is now routinely applied to iPSC-derived neurons to reveal primary cellular phenotypes in NDDs [34] [36].
Table 2: Common Patch Clamp Configurations and Their Applications
| Configuration | Description | Key Measurable Parameters |
|---|---|---|
| Whole-Cell | The membrane patch under the pipette is ruptured, providing electrical and physical access to the intracellular space. | Action potential properties (threshold, amplitude, frequency), postsynaptic potentials (EPSPs/IPSPs), membrane properties (capacitance, resistance) [34]. |
| Cell-Attached | The pipette forms a high-resistance seal on the cell membrane, but the patch is not ruptured. | Action potential firing patterns in a minimally perturbed cell, without dialysis of the intracellular contents [34]. |
| Voltage-Clamp | The amplifier actively clamps the cell's membrane voltage to a set command potential and measures the current required to maintain it. | Ionic currents through voltage-gated or ligand-gated channels (e.g., Na+, K+, Ca2+ currents); synaptic currents (AMPA, NMDA, GABAA receptor-mediated) [34]. |
| Current-Clamp | The amplifier injects a set current and records the resulting changes in membrane voltage. | Action potential generation and waveform, synaptic integration, and intrinsic excitability [34]. |
Experimental Protocol for iPSC-Derived Neurons:
- Cell Preparation: Plate iPSC-derived neurons on glass coverslips coated with appropriate substrates (e.g., poly-D-lysine, laminin). Recordings are typically performed after 4-8 weeks of differentiation to allow for sufficient maturation and synaptogenesis.
- Solution Preparation: Prepare an artificial cerebrospinal fluid (aCSF) for the bath solution, containing (in mM): 125 NaCl, 2.5 KCl, 2 CaCl₂, 1 MgCl₂, 1.25 NaH₂PO₄, 26 NaHCO₃, and 25 glucose, saturated with 95% O₂/5% CO₂ (pH 7.4). The internal pipette solution for whole-cell recordings typically contains (in mM): 130 K-gluconate, 6 KCl, 4 ATP-Mg, 0.3 GTP, 10 HEPES, and 0.2 EGTA.
- Recording Setup: Place the coverslip with neurons in a recording chamber continuously perfused with oxygenated aCSF. Visualize neurons using an upright microscope with differential interference contrast (DIC) optics.
- Electrode and Sealing: Pull borosilicate glass capillaries to fabricate recording pipettes with a resistance of 3-6 MΩ. Position the pipette onto a neuron using a micromanipulator and apply gentle suction to achieve a gigaohm (GΩ) seal.
- Whole-Cell Access: For whole-cell configuration, apply additional brief suction or a voltage zap to rupture the membrane patch. Compensate for series resistance and cell capacitance.
- Data Acquisition: Perform recordings in voltage-clamp or current-clamp mode. Apply specific pharmacological agents (e.g., TTX to block Na+ channels, CNQX/APV to block glutamate receptors) to isolate specific currents or synaptic inputs.
Multi-Electrode Array (MEA) Electrophysiology
MEA recordings provide a non-invasive, label-free method for long-term monitoring of network-level activity from populations of neurons, enabling larger sample sizes and longitudinal studies [34] [35]. This is particularly useful for capturing emergent properties in iPSC-derived neural networks and 3D organoids.
Experimental Protocol:
- Plate Preparation: Seed iPSC-derived neurons or neural progenitor cells directly onto MEA plates pre-coated with adhesion factors. For 3D models, place organoids or spheroids onto the electrode grid using specialized plates with guiding funnels [35].
- Culture Maintenance: Maintain the cultures in the MEA plates within a standard cell culture incubator, allowing networks to form and mature over weeks.
- System Setup: Place the MEA plate into the recording station, which provides environmental control (temperature, CO₂). The system includes a headstage for signal amplification and a computer for data acquisition [35].
- Data Recording: Record spontaneous electrical activity for specified durations (e.g., 10-30 minutes). The system simultaneously captures signals from all electrodes, detecting individual neuronal spikes and lower-frequency local field potentials [39] [35].
- Data Analysis: Use integrated software to extract key metrics, including:
- Mean Firing Rate: The average number of spikes per second across the network.
- Burst Detection: Identification of periods of high-frequency, synchronous spiking. Parameters include burst duration, inter-burst interval, and spikes per burst [39].
- Network Synchrony: Measures of correlated activity across multiple electrodes, reflecting the functional connectivity of the network.
A key advantage of MEA is its ability to reveal that the membrane potential of even a single neuron can serve as a highly sensitive probe for the cooperative burst activities of the entire network, a correlation that holds even for neurons located millimeters away from the electrode array [39].
Intracellular Calcium Imaging
Calcium imaging leverages the fact action potentials and synaptic activity are accompanied by rapid influxes of calcium ions into the cytoplasm. By measuring these changes, researchers can infer neuronal activation and characterize signaling phenotypes [37].
Experimental Protocol:
- Dye Loading or Genetic Encoding: Load cells with a synthetic, cell-permeable fluorescent calcium indicator (e.g., Cal-520 AM, Fluo-4 AM). Alternatively, use iPSC lines genetically engineered to express GCaMP, a genetically encoded calcium indicator.
- Experimental Setup: Place the cultured neurons or organoids on a microscope stage equipped with an appropriate light source (e.g., LED or laser) and a high-sensitivity camera (e.g., sCMOS). Maintain environmental control.
- Image Acquisition: Acquire time-lapse images at a high frame rate (10-100 Hz). For wide-field calcium imaging, a single wavelength is excited, and the emission is collected [38].
- Stimulation (Optional): Apply specific agonists or neuronal stimuli (e.g., high K+ solution, glutamate) to evoke calcium transients and probe the responsiveness of specific receptor pathways [37].
- Data Processing and Analysis:
- Region of Interest (ROI) Definition: Manually or automatically define ROIs around individual neuronal somas.
- Fluorescence Trace Extraction: For each ROI, extract the fluorescence intensity (F) over time.
- Calculation of (\Delta F/F): Normalize the trace by the baseline fluorescence ((F_0)) to obtain (\Delta F/F), which represents the relative change in intracellular calcium.
- Phenotype Characterization: Analyze transients for amplitude, kinetics (rise and decay time), frequency, and spatial propagation [37]. This data can be used for functional connectivity analysis or high-throughput screening [37] [18].
Calcium Imaging Workflow
Signaling Pathways and Functional Perturbations in NDDs
In iPSC-based NDD models, these functional assays are used to probe specific, dysregulated signaling pathways. A prominent example is the metabotropic glutamate receptor (mGluR) pathway in Fragile X Syndrome (FXS). In FXS-iPSC-derived neural progenitor cells, exaggerated mGluR signaling leads to increased calcium influx through calcium-permeable AMPA receptors, which contributes to impaired neuronal differentiation [20]. Calcium imaging would directly capture this hyper-responsive (Ca^{2+}) release upon mGluR agonist application, while patch clamp could isolate the specific mGluR-mediated currents and the properties of the AMPA receptors involved.
mGluR Pathway Dysregulation in FXS
The Scientist's Toolkit: Essential Research Reagents and Materials
Successful functional phenotyping relies on a suite of specialized reagents and instruments.
Table 3: Essential Materials for Functional Phenotyping of iPSC-Derived Neurons
| Category | Item | Function / Application |
|---|---|---|
| Cell Sources | Patient-derived iPSCs [20] | Provides a disease-relevant genetic background for modeling NDDs. |
| Isogenic control iPSCs [4] | CRISPR/Cas9-corrected lines from the same patient, serving as the ideal negative control to confirm phenotype-genotype links. | |
| Differentiation & Culture | Neural Induction Kits | Directs the differentiation of iPSCs into neural progenitor cells (NPCs) and specific neuronal subtypes. |
| Growth Factors (BDNF, GDNF, NT-3) | Promotes neuronal maturation, survival, and synaptic development in vitro. | |
| Functional Assay Reagents | Synthetic Calcium Indicators (Fluo-4 AM, Cal-520 AM) [37] | Cell-permeable dyes that fluoresce upon binding intracellular calcium for imaging. |
| Pharmacological Agonists/Antagonists (e.g., CNQX, APV, Bicuculline) | Tool compounds to selectively activate or block specific receptors and ion channels during patch clamp or MEA experiments. | |
| Key Instruments | Patch Clamp Amplifier & Micromanipulators [34] | Core hardware for high-resolution intracellular recording. |
| Multi-Electrode Array (MEA) System [35] | Platform for non-invasive, long-term network electrophysiology from 6- to 96-well plates. | |
| Inverted/Upright Microscope with EMCCD/sCMOS Camera | Essential for patch clamp visualization and high-sensitivity calcium imaging. | |
| Data Analysis | Analysis Software (e.g., Clampfit, Axograph, Neuroexplorer) | Specialized software for processing and analyzing electrophysiological and calcium imaging data. |
Patch clamp electrophysiology, multi-electrode arrays, and calcium imaging form a powerful, complementary toolkit for functional phenotyping in iPSC models of neurodevelopmental disorders. Patch clamp offers unrivaled resolution for dissecting mechanistic defects at the single-channel and single-synapse level. MEAs provide a macroscopic view of emergent network properties and are ideal for longitudinal studies and higher-throughput screening. Calcium imaging bridges the gap, allowing for the mapping of activity across large populations of cells with good spatial resolution. The convergence of these techniques with patient-specific iPSC models is rapidly advancing our understanding of the pathophysiological mechanisms underlying rare NDDs like Fragile X syndrome, Rett syndrome, and Dup15q, thereby illuminating new paths for therapeutic development.
The advent of induced pluripotent stem cell (iPSC) technology has revolutionized the study of human neurodevelopmental disorders by providing unprecedented access to patient-specific neural cells. This technical guide details the application of iPSC models to investigate two distinct genetic neurological conditions: NRXN1 deletions in schizophrenia (SCZ) and NTRK1 mutations in Hereditary Sensory and Autonomic Neuropathy Type IV (HSAN IV). These case studies exemplify how iPSC-derived neural models can bridge genetic findings to cellular and molecular disease mechanisms, offering powerful platforms for drug discovery and pathway analysis within the broader context of neurodevelopmental disorder research.
NRXN1 in Schizophrenia: iPSC Modeling of Synaptic Dysfunction
Background and Genetic Association
NRXN1, located on chromosome 2p16.3, encodes the presynaptic cell adhesion molecule neurexin-1. Heterozygous deletions in NRXN1 are strongly associated with multiple neuropsychiatric disorders, including schizophrenia (odds ratio = 14.4), autism spectrum disorder (odds ratio = 14.9), and epilepsy (odds ratio = 9.91) [40]. Despite high heritability, individuals with NRXN1 deletions display remarkable phenotypic variability, ranging from neurotypical presentation to severe neurodevelopmental disorders, suggesting significant contributions from genetic background and modifier genes [4].
Key Methodologies and Experimental Protocols
Village Editing and Multi-Donor iPSC Platforms
A novel "village editing" approach has been developed to study NRXN1 mutations across diverse genetic backgrounds. This method involves CRISPR/Cas9-mediated gene editing in a cell village format, enabling parallel generation of NRXN1 knockouts across multiple donor iPSC lines [4].
Detailed Protocol:
- Select iPSC lines from 15+ donors with varying polygenic risk scores for schizophrenia
- Perform CRISPR/Cas9 editing in pooled iPSC cultures (cell villages) targeting NRXN1
- Recover and clone edited cells: ~28.4% achieve homozygous deletion, ~33.1% heterozygous deletion
- Differentiate edited iPSCs to cortical excitatory neurons using dual SMAD inhibition
- Co-culture with mouse glia for 28+ days to enhance maturation
- Analyze transcriptomes via RNA sequencing and functional properties via calcium imaging and electrophysiology [4] [41]
Forebrain Organoid Modeling
3D forebrain organoids provide more complex modeling of cortical development:
Differentiation Protocol:
- Pattern hPSCs toward dorsal forebrain fate using SMAD inhibition and WNT activation
- Maintain in 3D Matrigel droplets with neuronal maturation media
- Collect samples at critical developmental timepoints: 3 weeks (progenitor expansion), 2 months (early neurogenesis), 3.5 months (synapse formation, astrogenesis)
- Validate organization: SOX2+ ventricular zones, HOPX+ outer radial glia, SATB2+/BCL11B+ cortical neurons
- Confirm synaptic maturation: SYNAPSIN+/SYNAPTOPHYSIN+ presynaptic, HOMER+ postsynaptic compartments [41]
Key Findings and Quantitative Data
Table 1: Quantitative Findings from NRXN1 iPSC Studies
| Experimental Measure | Finding | Significance | Source |
|---|---|---|---|
| NRXN1α isoform reduction in 5'-NRXN1+/− neurons | Significant decrease (P = 0.001) | Promoter-specific effects | [40] |
| NRXN1β isoform increase in 5'-NRXN1+/− neurons | Significant increase (P = 0.001) | Compensatory mechanism | [40] |
| Altered synaptic function | Impaired neurotransmitter release | Direct functional consequence | [42] |
| CASK protein levels | Increased in NRXN1 mutants | Alternative synaptic pathway activation | [42] |
| Single-cell transcriptomic perturbations | Enhanced in SCZ vs. isogenic background | Genetic background modulation | [41] |
Signaling Pathways and Molecular Mechanisms
NRXN1 Pathophysiology Network
NTRK1 in HSAN IV: Modeling Sensory Neuropathy
Background and Genetic Basis
Hereditary Sensory and Autonomic Neuropathy Type IV (HSAN IV) is a rare autosomal recessive disorder caused by mutations in the NTRK1 gene, which encodes the Tropomyosin Receptor Kinase A (TRKA). Clinical manifestations include congenital insensitivity to pain, anhidrosis (inability to sweat), and intellectual disability [4] [43]. The TRKA receptor serves as the primary signaling receptor for Nerve Growth Factor (NGF), essential for the development and survival of nociceptive neurons and autonomic nervous system components [44].
Key Methodologies and Experimental Protocols
DRG Organoid Modeling
Dorsal Root Ganglia (DRG) organoids have been established to model human sensory neuropathy:
Differentiation Protocol:
- Generate iPSCs from HSAN IV patients' urine samples
- Correct mutations in isogenic controls using CRISPR-based gene editing
- Differentiate iPSCs into 3D DRG organoids via neural crest induction
- Analyze developmental stages for neuronal and glial differentiation
- Assess markers: ISLET1+/BRN3A+ sensory neurons, TRKA/PV/CGRP+ nociceptors, FABP7+ glial cells [4]
Comprehensive Functional Analysis of NTRK1 Mutations
Detailed Functional Assay Protocol:
- Express seven NTRK1 missense mutations (e.g., p.G517E, p.L657P, p.C752S) in appropriate cell lines
- Assess TRKA glycosylation states via Western blotting
- Quantify cell surface expression using flow cytometry
- Measure NGF-stimulated autophosphorylation and Y496 phosphorylation
- Evaluate downstream PLCγ activation and neurite outgrowth in neuronal models [44]
Key Findings and Quantitative Data
Table 2: Experimental Findings from HSAN IV Modeling
| Experimental Measure | Finding | Significance | Source |
|---|---|---|---|
| Sensory neuron reduction in patient DRG organoids | Marked reduction in ISLET1+/BRN3A+ cells | Developmental deficit | [4] |
| Lineage switching | Neuronal to glial fate shift | Novel disease mechanism | [4] |
| Mature sensory marker expression | TRKA, TRKC, TRPV, CGRP significantly reduced | Functional impairment | [4] |
| Premature gliogenesis | Significant FABP7 upregulation | Altered developmental timing | [4] |
| Mutation functional diversity | 1 fully functional, 1 partially active, 4 null mutations | Complex pathogenicity | [44] |
Signaling Pathways and Molecular Mechanisms
NTRK1 Signaling Disruption in HSAN IV
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Research Reagents for iPSC Modeling of Neurodevelopmental Disorders
| Reagent/Category | Specific Examples | Function/Application | Source |
|---|---|---|---|
| Gene Editing Tools | CRISPR/Cas9, AAV-based recombination | Introduction of patient-specific mutations, isogenic control generation | [4] [42] |
| Differentiation Kits | Dual SMAD inhibitors, NGN2, ASCL1/DLX2 | Directed differentiation to neural lineages | [40] [41] |
| Cell Culture Matrices | Matrigel, Laminin, Poly-ornithine | 3D organoid formation, neuronal adhesion | [41] |
| Neuronal Markers | MAP2, SOX2, NESTIN, ISLET1, BRN3A | Cell type identification, differentiation validation | [4] [40] |
| Synaptic Markers | SYNAPSIN, SYNAPTOPHYSIN, HOMER, CASK | Synapse formation and function assessment | [41] [42] |
| Functional Assays | Calcium imaging, MEA, autophosphorylation assays | Neuronal activity and signaling measurement | [41] [44] |
| Omics Technologies | scRNA-seq, long-read RNA sequencing, spatial transcriptomics | Molecular profiling, isoform analysis | [40] [41] |
Comparative Analysis and Future Directions
These case studies demonstrate how patient-derived iPSC models provide critical insights into disorder-specific pathophysiological mechanisms. While NRXN1 deletions primarily disrupt synaptic organization and neuronal network activity, NTRK1 mutations fundamentally alter sensory neuron development and survival. Both models highlight the importance of genetic background in modulating phenotypic outcomes and the necessity of isogenic controls for proper interpretation of disease-associated variants.
Future methodological developments will focus on enhancing organoid complexity through assembloid approaches, improving functional maturation of neuronal cultures, and integrating multi-omics datasets to build comprehensive molecular networks. These advanced models will accelerate the identification of therapeutic targets and facilitate personalized medicine approaches for neurodevelopmental disorders.
The integration of Organ-on-a-Chip (OoC) technology with advanced cell culture models represents a transformative approach for investigating human physiology and disease. This whitepaper details how these microphysiological systems, particularly when combined with induced pluripotent stem cell (iPSC)-derived models, are advancing the study of neurodevelopmental disorders. OoCs provide a controlled microenvironment to simulate tissue-specific functions, mechanical cues, and multi-organ interactions that are essential for understanding complex disease mechanisms and accelerating drug discovery. This guide provides a technical framework for developing these sophisticated models, with a specific focus on neurological applications.
Organ-on-a-Chip (OoC) systems are microfluidic devices lined with living human cells that are designed to recapitulate the minimal functional unit of an organ [45]. These systems have emerged as a powerful alternative to traditional two-dimensional (2D) cell cultures and animal models, which often fail to predict human therapeutic responses due to species-specific differences and an inability to mimic the human tissue microenvironment [46] [47]. The foundational concept of reproducing organic and physiological functions on a chip was first published in 2004, with the term "Organ-on-a-Chip" being formally adopted in 2010 [46].
The core value of OoC technology lies in its ability to mimic three key aspects of organ physiology:
- Barrier functions composed of multiple tissues and vasculature
- Parenchymal organization with different tissue substructures within the same organ
- Interaction between different organs [46]
For neurodevelopmental disorder research, OoCs offer a unique platform to study human-specific developmental processes, neural network formation, and the functional impact of genetic variations within a controlled yet physiologically relevant context.
Table 1: Comparison of Preclinical Model Systems
| Characteristics | Animal Models | 2D Cell Cultures | Organ-on-Chip |
|---|---|---|---|
| Target tissue representativeness | Low | Low | High |
| Ethical implications | Medium | Low | Low |
| Associated costs | High | Low | Low |
| Viability maintenance | Medium | High | High |
| High-throughput analysis aptitude | Low | High | Medium |
| Full body representativeness | High | Low | Medium |
| Physiological microenvironment | Limited | Poor | High |
Core Components and Design Principles
Engineering Considerations
Developing a functional OoC requires an interdisciplinary approach combining microengineering, cell biology, and biomaterials science [48]. The initial design phase must identify the scientific question and define the minimal functional unit of the target organ to be recapitulated.
Material Selection: The choice of chip material depends on the intended application. Polydimethylsiloxane (PDMS) remains widely used due to its ease of fabrication, gas permeability, and optical clarity; however, its tendency to absorb small molecules can be problematic for pharmacokinetic studies [48]. Thermoplastics offer an alternative with minimal absorption but lower gas permeability, potentially requiring integrated oxygenation strategies. Increasingly, hybrid material chips are being developed to balance these properties.
Sensor Integration: The transparent nature of most microfluidic platforms enables excellent optical accessibility, while integrated sensors allow real-time monitoring of cell culture conditions and biological function [48]. For neural models, this may include electrical activity sensors or metabolic sensors to monitor neuronal function.
OoC Development Workflow
Biological Components
Cell Sources: OoCs can utilize multiple cell sources, each with distinct advantages:
- Immortalized cell lines: Cost-effective and standardized, but often lack critical phenotypical characteristics
- Primary cells: Offer mature phenotypes but have limited availability
- iPSCs: Provide autologous, patient-specific cells with potential for standardization [48]
For neurodevelopmental disease modeling, iPSCs are particularly valuable as they allow the generation of patient-specific neural tissues while maintaining the genetic background of the donor.
Biomaterials and Scaffolds: The extracellular matrix (ECM) provides critical physical and biochemical cues that guide tissue assembly and functional maturation. Both scaffold-based (typically hydrogels) and scaffold-free approaches are employed, with the choice dependent on the specific tissue requirements and experimental timeframe [48].
Table 2: Research Reagent Solutions for OoC Development
| Category | Specific Examples | Function/Application |
|---|---|---|
| Chip Materials | PDMS, Thermoplastics (PMMA, PS), Hybrid materials | Structural foundation with specific absorption and permeability properties |
| Cell Sources | iPSCs, Primary neural cells, Immortalized lines | Provide biological functionality with varying physiological relevance |
| Biomaterials/Scaffolds | Natural hydrogels (Matrigel, collagen), Synthetic PEG-based hydrogels | Mimic extracellular matrix to support 3D tissue architecture |
| Sensors/Actuators | Optical oxygen sensors, TEER electrodes, Mechanical stretchers | Monitor and manipulate microenvironment in real-time |
| Perfusion Media | Specialized neuronal culture media, Blood-brain barrier media | Provide nutritional support and physiological signaling molecules |
Integration with iPSC Technology for Neurological Disease Modeling
The combination of OoC with iPSC technology has opened new avenues for modeling neurological disorders. This approach enables researchers to create patient-specific neural models that recapitulate key aspects of human brain development and disease pathology.
Case Studies in Neurodegeneration: Research led by Dr. Clive Svendsen at Cedars-Sinai has demonstrated how OoC technology can be used to build more physiological models of neurodegeneration. By generating chips with neural tissue and a blood-brain-barrier (BBB)-like structure from iPSCs, his team has developed models for Amyotrophic Lateral Sclerosis (ALS), Parkinson's disease, and frontotemporal dementia (FTD) [49]. These models enable the prediction of pathology and allow interactive studies to predict drug effects in a human-relevant system.
Modeling Neurodevelopmental Disorders: For neurodevelopmental disorders, the integration of OoC with iPSCs is particularly powerful. Patient-derived iPSCs can be differentiated into various neural cell types (neurons, astrocytes, oligodendrocytes) and assembled into 3D structures that mimic the developing brain. When cultured in OoC platforms, these models can be subjected to precise environmental control and mechanical stimulation that better mimics the in vivo microenvironment.
iPSC to OoC Model Pipeline
Experimental Protocols and Methodologies
OoC Fabrication Protocol
Materials:
- PDMS base and curing agent (typically 10:1 ratio)
- SU-8 photoresist for master mold fabrication
- Silicon wafers
- Plasma cleaner for bonding
- Sterile culture hood and equipment
Procedure:
- Master Mold Fabrication: Create the master mold using photolithography with SU-8 photoresist on silicon wafers.
- PDMS Casting: Mix PDMS base and curing agent, degas under vacuum, and pour onto the master mold.
- Curing and Bonding: Cure at 65°C for 4 hours, peel off from mold, and create access ports using biopsy punches.
- Sterilization: Bond PDMS to glass substrate using oxygen plasma treatment, then sterilize by autoclaving or UV irradiation.
- Surface Treatment: If necessary, treat channels with extracellular matrix proteins (e.g., fibronectin, collagen) to enhance cell adhesion.
iPSC Differentiation and OoC Seeding
Neural Induction Protocol:
- iPSC Maintenance: Culture iPSCs in feeder-free conditions using defined essential medium.
- Neural Induction: Transition cells to neural induction medium containing dual SMAD inhibitors (dorsomorphin and SB431542) for 10-14 days.
- Neural Progenitor Expansion: Passage cells and expand in neural progenitor medium containing FGF2 and EGF.
- OoC Seeding: Harvest neural progenitors and seed into the OoC device at high density (10-50 million cells/mL).
- Maturation: Maintain chips under continuous perfusion with neural differentiation medium for 4-8 weeks to allow maturation and network formation.
Functional Assessment Methods
Barrier Integrity Assessment:
- Transepithelial/Transendothelial Electrical Resistance (TEER): Use integrated electrodes to measure electrical resistance across barriers.
- Permeability Assays: Introduce fluorescent dextrans of varying sizes and measure flux across the barrier.
Functional Neural Assessment:
- Calcium Imaging: Use Fluo-4 or similar calcium indicators to monitor neural activity and network synchronization.
- Multi-electrode Arrays (MEAs): Record electrical activity from neuronal networks.
- Immunocytochemistry: Fix and stain for neural markers (MAP2, β-III-tubulin), synaptic markers (synapsin, PSD-95), and glial markers (GFAP).
Table 3: Quantitative Parameters for OoC Model Validation
| Parameter | Traditional 2D Culture | Organ-on-Chip | In Vivo Human Reference |
|---|---|---|---|
| Barrier Integrity (TEER, Ω×cm²) | 50-200 | 600-1500 | 1000-2000 (BBB) |
| Metabolic Activity (per cell) | Low | High, sustained | High |
| Gene Expression Profile | Fetal-like | Adult-like | Adult |
| Drug Sensitivity | Often hyper-sensitive | Physiologically relevant | Clinical response |
| Cellular Diversity | Limited | High, controlled | High |
Applications in Neurodevelopmental Disorder Research
OoC technology enables specific applications that are particularly relevant to neurodevelopmental disorder research:
Disease Modeling with Patient-specific Cells: The ability to incorporate patient-derived iPSCs into OoCs allows researchers to create personalized models of neurodevelopmental disorders. For example, models of spinal muscular atrophy (SMA) have successfully replicated disease-specific features including motor neuron defects and aberrant neural stem cell differentiation [50].
Blood-Brain Barrier Modeling: Neurological OoCs can incorporate a functional blood-brain barrier (BBB) by co-culturing brain microvascular endothelial cells with pericytes and astrocytes. This allows researchers to study BBB breakdown in disease states and evaluate drug penetration across the BBB [49].
Multi-organ Interaction Studies: For neurodevelopmental disorders with systemic manifestations, multi-OoC systems can be fluidically linked to study gut-brain, liver-brain, or other organ-organ interactions. These systems can recapitulate interdependent organ functions and systemic drug metabolism [46] [47].
Future Perspectives and Challenges
While OoC technology holds tremendous promise, several challenges must be addressed for broader adoption:
Standardization and Scaling: Current OoC platforms vary significantly between laboratories, making direct comparison of results challenging. Efforts are underway to develop standardized platforms and operating procedures to enhance reproducibility [48].
Complexity Management: As models become more sophisticated, balancing physiological relevance with practical usability becomes increasingly important. The goal should be systems that are "as simple as possible, but as complex as necessary" to answer the specific research question [48].
Regulatory Acceptance: As OoC technology matures, regulatory agencies are increasingly recognizing its potential. In 2022, the U.S. FDA announced that animal testing would no longer be required prior to clinical trials, opening the door for alternative models like OoCs [50]. Further validation studies are needed to fully establish OoCs as predictive tools for drug development.
The integration of single-cell technologies (transcriptomics, proteomics) with OoCs will provide unprecedented resolution to understand how specific drugs interact with different cell types within the chip [49]. This multi-modal approach will significantly advance our understanding of neurodevelopmental disorders and accelerate the development of effective therapeutics.
Navigating iPSC Challenges: Solutions for Standardization, Maturation, and Scalability in Neural Modeling
The maturation state of neurons is a critical determinant of brain circuit function and stability. Within the context of induced pluripotent stem cell (iPSC) modeling of neurodevelopmental disorder (NDD) mechanisms, a pervasive observation is the prevalence of cellular immaturity—a state where neurons exhibit molecular, morphological, and physiological characteristics reminiscent of earlier developmental stages. This whitepaper synthesizes current research to detail the mechanistic basis of neuronal immaturity and provides a strategic framework for experimental approaches aimed at enhancing neuronal aging and functional maturation. We explore the roles of key signaling pathways, present advanced iPSC-derived model systems, and provide a detailed "Scientist's Toolkit" of reagents and methodologies. By integrating recent findings on opposing "hyper-maturity" states, this guide aims to equip researchers with the tools to precisely modulate neuronal maturation, thereby advancing the study of NDD pathogenesis and therapeutic development.
The human brain is characterized by a dynamic balance between neuronal stability and plasticity. A deviation from typical maturational trajectories, resulting in arrested development or a pseudo-immature state, is increasingly implicated in the pathophysiology of diverse neurodevelopmental and neuropsychiatric conditions [51] [52]. In the realm of iPSC-based disease modeling, a common and significant phenotype is the observation that patient-derived neurons often display reduced complexity, altered electrophysiological properties, and an expression profile skewed towards immaturity even after extended culture periods [18] [53]. This cellular immaturity is not merely a technical artifact but is believed to reflect intrinsic pathophysiological mechanisms, potentially contributing to the cognitive deficits and circuit dysfunctions observed in disorders such as autism spectrum disorder (ASD), intellectual disability (ID), and schizophrenia [52] [53].
The translational importance of studying brain plasticity, including maturational states, is paramount. Given that most neurological disorders remain incurable, understanding the exceptions to neural stability offers potential targets for preventive and therapeutic approaches [51]. The discovery that maturational states of brain cells can change plastically in response to genetic and environmental insults provides a foundation for intervention [52]. However, recent evidence complicates this picture, revealing that the opposite phenomenon—hyper-maturity and accelerated aging in the hippocampus—is also associated with neuropsychiatric phenotypes, particularly anxiety-like behavior [52] [54]. This underscores that a precise, homeostatic balance in neuronal maturation is critical for healthy brain function, and iPSC models provide the essential platform for dissecting these complex dynamics within a human genetic context.
Molecular Mechanisms of Neuronal Maturation
The transition from an immature to a mature neuron involves a coordinated sequence of transcriptional, epigenetic, and cellular changes. Disruptions in this finely tuned process can lead to the persistent immaturity observed in NDD models.
Key Molecular Markers of Maturation
Tracking neuronal maturation requires a firm understanding of stage-specific biomarkers. The following table summarizes key molecules used to assess neuronal maturity.
Table 1: Key Molecular Markers for Assessing Neuronal Maturity
| Marker | Full Name | Expression in Immature Neurons | Expression in Mature Neurons | Primary Function |
|---|---|---|---|---|
| DCX | Doublecortin | High | Low/Virtually Absent | Microtubule-associated protein; critical for neuronal migration and differentiation [51] |
| PSA-NCAM | Polysialylated Neural Cell Adhesion Molecule | High | Low | Membrane-bound glycoprotein; modulates cell-cell interactions and structural plasticity [51] |
| TBR1 | T-Box Brain Transcription Factor 1 | Low | High | Critical transcription factor for cortical identity and synaptic function [53] |
| SYNAPSIN I | Synapsin I | Low | High | Presynaptic protein; regulates neurotransmitter release and synaptogenesis [55] |
| GRIN2B | Glutamate Ionotropic Receptor NMDA Type Subunit 2B | High | Low (in some contexts) | NMDA receptor subunit; subunit composition shifts (e.g., from GluN2B to GluN2A) during maturation [54] |
| CAMK2A | Calcium/Calmodulin Dependent Protein Kinase II Alpha | Low | High | Synaptic signaling kinase; its upregulation is associated with hyper-maturity [54] |
Signaling Pathways Governing Maturation
Several evolutionarily conserved signaling pathways act as master regulators of the neuronal maturation process. Targeting these pathways offers strategic leverage to modulate the maturational state of iPSC-derived neurons.
- mTORC1 Signaling: The mechanistic target of rapamycin complex 1 (mTORC1) is a central integrator of cellular signals that promotes protein synthesis, dendritic growth, and spinogenesis. Its activation is a common downstream effect of rapid-acting antidepressants like ketamine and its metabolite (2R,6R)-HNK, as well as the glycoprotein Reelin [55]. In Tuberous Sclerosis Complex (TSC), mTORC1 hyperactivation leads to neuronal hyperexcitability, demonstrating that its activity must be precisely tuned for normal maturation [18].
- Reelin Signaling: The large extracellular glycoprotein Reelin, secreted by Cajal-Retzius cells during development and specific interneuron populations in adulthood, governs neuronal positioning, dendritic arborization, and synaptic plasticity. Reelin is frequently downregulated in depression, and its restoration has demonstrated antidepressant-like effects and promotes synaptic maturation by modulating mTORC1 activity and downstream synaptic protein expression (e.g., GluA1, PSD-95) [55].
- Activity-Dependent Pathways: Neuronal activity is a critical driver of maturation. Suppression of activity can maintain immaturity, while specific patterns of excitation can promote maturation. Chronic stress or corticosterone exposure can induce a hyper-mature/aged transcriptomic profile, linking environmental factors to accelerated maturational decline [52] [54].
The following diagram illustrates the interplay between these key pathways in regulating neuronal maturation.
Figure 1: Key Signaling Pathways in Neuronal Maturation. External cues like Reelin and neuronal activity through NMDARs converge on mTORC1 signaling, which upregulates synaptic proteins (SynProt) and promotes structural plasticity (StructPlast), collectively driving neuronal maturation.
Experimental Models for Studying Neuronal Maturation
Moving beyond traditional two-dimensional (2D) cultures, advanced iPSC-derived models now offer unprecedented ability to recapitulate the complexity of human brain development.
Advanced iPSC-Derived Model Systems
- 2D Monolayer Cultures: Patient-specific iPSCs are differentiated into relatively pure populations of cortical excitatory or inhibitory neurons. These systems are ideal for high-throughput drug screening and detailed electrophysiological characterization (e.g., multielectrode arrays) [18] [55]. A key study on treatment-resistant depression (TRD) used this model to show that (2R,6R)-HNK and Reelin similarly modulate synaptic protein expression in TRD patient-derived neurons [55].
- 3D Brain Organoids: These self-organizing, three-dimensional structures mimic the cellular composition, architectural complexity, and cell-cell interactions of the developing human brain more accurately than 2D models [30]. Region-specific organoids (e.g., cortical, striatal) provide highly reproducible systems for studying cell-intrinsic maturation pathways within a specific brain region.
- Assembloids: To model circuit-level maturation and integration, organoids from different brain regions (e.g., cortical and striatal) can be fused to create assembloids. These models enable the study of long-range axonal projection, synaptic connectivity, and interneuron migration—all critical processes for functional maturation [4] [30]. For example, dorsal-ventral assembloids have been used to model the prolonged migration of interneurons observed in the postnatal human brain, a process vulnerable to insult [4].
Quantifying Maturation in Experimental Models
A multi-parametric approach is essential for accurately assessing the maturity of iPSC-derived neuronal models. The table below outlines key metrics.
Table 2: Metrics for Quantifying Neuronal Maturation in iPSC-Derived Models
| Metric Category | Specific Assay/Readout | What It Measures |
|---|---|---|
| Gene Expression | RNA Sequencing (Bulk/Single-cell) | Transcriptomic profile, maturity index, pathway enrichment (e.g., synaptic genes) [52] [55] |
| Protein Expression | Immunocytochemistry / Western Blot | Levels of maturity markers (TBR1, SYNAPSIN) vs. immaturity markers (DCX, PSA-NCAM) [51] [55] |
| Morphological | Dendritic Arborization Analysis | Complexity (Sholl analysis), total dendritic length, spine density and morphology |
| Functional | Patch Clamp / Multielectrode Array (MEA) | Action potential properties, spontaneous postsynaptic currents, network bursting activity [18] |
| Metabolic | Metabolomic Profiling (e.g., MetaboLINK) | Stage-specific metabolic programs involving amino acids, lipids, and energy utilization [18] |
The Scientist's Toolkit: Research Reagent Solutions
This section details essential reagents and tools for designing experiments aimed at modulating and assessing neuronal maturation.
Table 3: Research Reagent Solutions for Studying Neuronal Maturation
| Reagent / Tool | Function/Mechanism | Example Application in Research |
|---|---|---|
| (2R,6R)-HNK | Ketamine metabolite; activates mTORC1 signaling without NMDAR blockade. | Increased expression of GluA1, PSD-95, and Synapsin I in iPSC-derived cortical neurons from TRD patients [55]. |
| Reelin Protein | Endogenous glycoprotein; activates mTORC1 and downstream synaptic signaling. | Rescued stress-induced synaptic deficits and demonstrated rapid antidepressant-like effects in models; applied to iPSC-derived neurons at 50 nM [55]. |
| CRISPR/Cas9 | Genome editing for isogenic control generation or gene knockout. | "Village editing" approach to create NRXN1 knockouts in iPSCs from 15 donors to study genetic background effects [4]. |
| Corticosterone | Glucocorticoid receptor agonist; models chronic stress. | Induced hippocampal hyper-maturity and accelerated aging transcriptomic signatures in mouse models [52]. |
| mTORC1 Inhibitors | Suppresses hyperactive mTOR signaling (e.g., Rapamycin). | Rescued hyperexcitability and abnormal morphology in TSC2−/− iPSC-derived neurons [18]. |
| DCX Antibody | Immunohistochemical marker for immature, newborn neurons. | Used to detect and quantify populations of immature neurons in human post-mortem tissue and organoid cultures [51] [56]. |
Experimental Protocols for Modulating Maturation
Protocol: Testing Pro-Maturation Compounds in 2D iPSC-Derived Neuronal Cultures
This protocol is adapted from studies investigating the effects of (2R,6R)-HNK and Reelin on human iPSC-derived neurons [55].
- Neuronal Differentiation: Differentiate control and patient-derived iPSCs into cortical excitatory neurons using a validated, multi-week protocol. Confirm neuronal identity (e.g., MAP2+ positivity) and the presence of synaptic markers (e.g., PSD-95, Synapsin I) via immunostaining at baseline.
- Compound Treatment: At a pre-defined maturation stage (e.g., Day 60-80 of differentiation), treat cultures with:
- Experimental Group 1: 1 µM (2R,6R)-HNK
- Experimental Group 2: 50 nM Reelin protein
- Vehicle Control Group
- Time-Course Harvesting: Harvest cells for analysis at multiple time points post-treatment (e.g., 1 hour, 24 hours) to capture rapid signaling changes and later transcriptional/protein-level effects.
- Downstream Analysis:
- Protein Analysis: Perform Western Blotting on cell lysates to quantify changes in synaptic proteins (GluA1, PSD-95, Synapsin I) and phosphorylation of key signaling nodes (e.g., p-ERK, Dab1).
- Transcriptomic Analysis: Conduct bulk RNA-sequencing on samples from the 1-hour time point to identify early gene expression changes and pathway enrichment (e.g., mTORC1 signaling).
- Functional Analysis: Perform live-cell calcium imaging or MEA on a parallel set of cultures to assess changes in network activity and synchronization.
Protocol: Generating and Validating a Hyper-Maturity Signature in Cortical Organoids
This protocol leverages recent findings on hippocampal hyper-maturity to investigate similar phenomena in forebrain models [52] [54].
- Organoid Generation: Generate dorsal forebrain organoids from iPSCs using a region-specific patterning protocol (e.g., using SMAD inhibitors and Wnt antagonists) [30].
- Induction of Hyper-Maturity:
- Chronic Stress Model: chronically expose organoids to 100 nM corticosterone for 40 days.
- Genetic Model: Introduce a genetic variant associated with a hyper-maturity phenotype (e.g., relevant to lysosomal storage disorders or SERT knockout) via CRISPR/Cas9.
- Transcriptomic Profiling: At the endpoint, perform RNA-sequencing on the organoids. Compare transcriptomes to:
- Age-matched control organoids.
- A published gene expression signature of hippocampal hyper-maturity and accelerated aging [52].
- Pathway and Behavioral Correlation:
- Conduct pathway enrichment analysis (e.g., using BaseSpace) to identify upregulated biological processes, with an expected enrichment for synaptic pathways (e.g., CAMK2A, GRIN2B).
- Correlate the in vitro hyper-maturity signature with in vivo behavioral data from corresponding mouse models, noting that hyper-maturity is associated with increased anxiety-like behaviors [54].
The workflow for this experimental approach is summarized below.
Figure 2: Experimental Workflow for Modeling Hyper-Maturity in Cortical Organoids. The process involves generating region-specific organoids, applying a genetic or stress-based intervention, and validating the resulting transcriptomic signature against known hyper-maturity profiles and associated behavioral phenotypes.
Therapeutic Implications and Future Perspectives
The strategic enhancement of neuronal maturation presents a promising therapeutic avenue for NDDs. Approaches that boost mTORC1 signaling, such as Reelin or (2R,6R)-HNK, could be explored to overcome immaturity in specific patient populations [55]. Conversely, the discovery of hyper-maturity as a pathological state underscores that the therapeutic goal is not simply to maximize maturation, but to restore a homeostatic balance [52] [54]. This necessitates patient stratification based on biomarkers of maturational state.
Future research must leverage multi-omics integration and machine learning to deconstruct the immense heterogeneity of maturational phenotypes across different genetic backgrounds [4] [18]. Furthermore, the development of more physiologically relevant models, such as vascularized organoids and assembloids that better recapitulate circuit-level maturation, will be critical for translating these strategies into effective therapies. The ultimate goal is to move the field toward predictive, patient-specific models that can guide the development of targeted interventions to correct aberrant maturational trajectories in neurodevelopmental disorders.
The application of induced pluripotent stem cells (iPSCs) in modeling neurodevelopmental disorders represents a transformative approach in neuroscience research. However, the immense potential of this technology is constrained by significant challenges in reproducibility across different laboratories and cell lines. Inconsistent differentiation outcomes and variable cellular quality can compromise disease modeling validity and drug screening reliability. This technical guide outlines standardized differentiation protocols and comprehensive quality control frameworks essential for ensuring reproducible results in neurodevelopmental disorder research. By establishing rigorous methodological standards and quantitative assessment criteria, researchers can enhance the reliability of iPSC-based models, thereby accelerating the identification of disease mechanisms and therapeutic targets.
Quality Control Metrics for iPSC and Neural Differentiation
Comprehensive QC Framework for Cerebral Organoids
For complex 3D models like cerebral cortical organoids, a systematic quality control (QC) framework is essential. A proposed methodology for 60-day cortical organoids evaluates five critical criteria using a standardized scoring system, each rated from 0 (low quality) to 5 (high quality) [57]:
Table: Quality Control Scoring Framework for 60-Day Cortical Organoids
| Criterion | Sub-Indices | Minimum Threshold Score | Assessment Method |
|---|---|---|---|
| Morphology | Surface integrity, compactness, border definition | 3 | Bright-field microscopy |
| Size & Growth Profile | Diameter, growth consistency | 3 | Microscopy with scale |
| Cellular Composition | Neural progenitors, neurons, glial cells | 4 | Immunostaining, flow cytometry |
| Cytoarchitectural Organization | Rosette formation, layered organization | 3 | Histology, immunostaining |
| Cytotoxicity | Cell death, necrosis | 4 | Viability dyes, LDH assay |
This hierarchical framework begins with non-invasive assessments (Criteria A-B) for initial screening, reserving in-depth analyses (Criteria C-E) for organoids passing initial evaluation. Implementation of this QC system has successfully discriminated organoid quality in validation studies using hydrogen peroxide-induced stress models [57].
Validated QC Tests for GMP-Compliant hiPSCs
For basic iPSC quality assurance, particularly for therapeutic applications, specific validated quality control tests should be implemented [58]:
- Residual Episomal Vector Testing: Accurate determination requires minimum input of 20,000 cells (120 ng genomic DNA), with screening recommended between passages 8-10 to avoid unnecessary rejection of lines
- Pluripotency Marker Assessment: Cutoff value set to expression of at least three individual markers on ≥75% of cells, using fluorescence minus one controls in multi-color flow cytometry panels to ensure accurate fluorescence spread
- Directed Differentiation Potential: Detection limit established at two of three positive lineage-specific markers for each of the three germ layers
- Genomic Stability Monitoring: Regular karyotyping at early passages (7-10) and every 10-15 passages during propagation, with STR profiling and mycoplasma testing conducted regularly [59]
Standardized Neuronal Differentiation Protocols
Optimized NGN2-Driven Glutamatergic Neuronal Differentiation
Transcription factor-based differentiation protocols, particularly Neurogenin-2 (NGN2) overexpression, significantly reduce heterogeneity and improve consistency across cell lines compared to extrinsic factor methods [60]. Key optimizations include:
NGN2 Differentiation Workflow
Critical improvements to standard NGN2 protocols address major sources of variability [60]:
- Pre-differentiation Genomic Screening: Employing high-resolution SNP Infinium arrays (560,000 probes) to identify genomic rearrangements undetectable by conventional karyotyping
- Homogeneous Population Selection: Using FACS sorting to isolate iPSCs with median, homogeneous GFP-NGN2 expression levels, reducing heterogeneity caused by variable transcription factor expression
- Intermediate Progenitor Banking: Incorporating a cryopreservation step for neuronal progenitors, enabling consistent starting material across multiple experiments
Essential Protocol Considerations
Successful implementation requires attention to several technical aspects:
- Culture Conditions: Use of chemically defined media such as Essential 8 (E8) under feeder-free conditions with appropriate extracellular matrices (Matrigel, Geltrex, or Laminin-521) [59]
- Passaging Methods: Employment of non-enzymatic, EDTA-based dissociation (Versene solution) to improve cell survival and replating efficiency [59]
- Cryopreservation Strategy: Large-batch freezing of characterized iPSCs at earlier passages to safeguard against contamination or phenotypic drift
The Scientist's Toolkit: Essential Research Reagents
Table: Essential Reagents for Standardized iPSC Neural Differentiation
| Reagent Category | Specific Examples | Function | Protocol Application |
|---|---|---|---|
| Reprogramming Vectors | Sendai virus vectors, episomal plasmids | Non-integrating delivery of reprogramming factors | iPSC generation [59] [60] |
| Culture Media | Essential 8 (E8), StemFlex | Chemically defined maintenance of pluripotency | iPSC expansion [59] |
| Extracellular Matrices | Matrigel, Geltrex, Laminin-521 | Support attachment and expansion | Feeder-free culture [59] |
| Differentiation Inducers | Doxycycline for Tet-On systems | Controlled transgene expression | NGN2-driven differentiation [60] |
| Selection Agents | Puromycin | Isolation of successfully transduced cells | Stable line generation [60] |
| Dissociation Reagents | Versene (EDTA solution) | Gentle, enzyme-free passaging | iPSC maintenance [59] |
| Detection Antibodies | Anti-NANOG, SOX2, OCT3/4, SSEA4 | Pluripotency marker assessment | Quality control [59] [60] |
Characterization of Functional Neuronal Maturation
Beyond morphological and molecular markers, functional assessment is crucial for validating neuronal maturation in neurodevelopmental disease models.
Functional Neuronal Assessment
Key functional metrics include [11]:
- Calcium Transients: Measured using GCaMP6s sensor, with reduced transient frequency indicating impaired neuronal activity (29.8% ± 0.7% in ASD models vs. 100% ± 1.3% in controls)
- Synaptic Transmission: Evaluated through spontaneous excitatory postsynaptic currents (sEPSC) frequency and amplitude
- Network Connectivity: Assessed using chemogenetic approaches (DREADDs) in co-culture systems or multi-electrode arrays (MEAs)
- miRNA Profiling: Identification of dysregulated miRNAs (hsa-let-7e-5p, hsa-miR-135b-5p, hsa-miR-16-5p, hsa-miR-27b-3p) in neuronal progenitor cells associated with neuronal signaling pathways [11]
Implications for Neurodevelopmental Disorder Research
Implementing standardized differentiation protocols and quality control metrics directly enhances the validity of disease mechanism studies in neurodevelopmental disorders like autism spectrum disorder (ASD). Functional analyses of iPSC-derived neurons from idiopathic ASD individuals reveal measurable deficits in neuronal activity, including reduced calcium transients and impaired synaptic neurotransmission [11]. These reproducible functional phenotypes provide a foundation for investigating molecular pathomechanisms and screening therapeutic compounds.
The relationship between genetic and epigenetic variation in iPSCs and their differentiated neuronal derivatives further underscores the importance of standardization. Studies demonstrate that epigenetic variation increases during differentiation, while the direct relationship with genetic variation becomes weaker [61]. This highlights the need for controlled differentiation conditions to minimize non-genetic sources of variability when modeling neurodevelopmental disorders with strong genetic components.
Standardized differentiation protocols and comprehensive quality control metrics are fundamental to achieving reproducibility in iPSC-based modeling of neurodevelopmental disorders. Implementation of the frameworks and methodologies outlined in this guide—from validated QC assays and optimized NGN2 differentiation to functional neuronal characterization—provides a pathway toward more reliable, comparable research outcomes. As the field advances toward therapeutic applications, these standardized approaches will be essential for generating clinically relevant insights into disease mechanisms and potential interventions.
The complexity of human brain development and the limited access to disease-relevant tissues present significant challenges in understanding neurodevelopmental disorders (NDDs). Induced pluripotent stem cell (iPSC) technology has emerged as a transformative platform for investigating conditions such as autism spectrum disorder (ASD) and schizophrenia (SCZ), which are now conceptualized as disorders of abnormal brain development that result in altered circuitry [62]. The diverse symptomatology of these conditions, combined with complex polygenic etiology, creates a major obstacle for mechanistic disease research [63]. While patient iPSC-derived neural cultures provide unprecedented opportunities to study disease-relevant cell types and investigate disease etiology, the technical hurdles surrounding their manufacturing remain substantial [63] [64].
The development costs for biopharmaceuticals are, on average, 11 times that of their small molecule counterparts, leading to product costs approximately 22 times higher [65]. This cost structure creates significant barriers for research and therapeutic development. Furthermore, iPSC manufacturing faces unique challenges in generating stable, well-characterized, pluripotent cells suitable for making differentiated cell types for research and clinical applications [64]. This technical guide examines the current state of automation, bioprocessing, and cost-reduction strategies specifically within the context of iPSC modeling of neurodevelopmental disorder mechanisms, providing researchers with actionable methodologies to enhance their experimental outcomes while managing resources effectively.
Technical Hurdles in iPSC Generation and Neural Differentiation
Intellectual Property and Regulatory Complexities
The iPSC intellectual property landscape for research and therapeutic applications is exceptionally complex, with numerous patents surrounding core technologies. A study from 2019 estimated that 1,516 patent applications for iPSC technologies were filed in the United States alone, with 895 in Japan, and 420 in Europe [64]. For researchers developing iPSC-based disease models, freedom to operate (FTO) requires careful consideration of multiple factors, including: choice of reprogramming factors, methods used to deliver reprogramming factors, choice of somatic donor cell type, culture conditions, and differentiation methods [64]. This complex patent landscape may necessitate pursuing alternative technology strategies to de-risk research programs, particularly when moving from basic research to clinical applications.
Donor Tissue Sourcing and Raw Material Considerations
Donor tissue sourcing represents a critical initial step in iPSC generation. Legal requirements mandate that donors must meet country-specific requirements for donation, procurement, and testing. When iPSC-derived models are intended for use in multiple jurisdictions, researchers must account for differences in regulatory requirements, which are not always consistent across regions [64]. Variations exist due to region-specific risk factors, such as restrictions on material from European origin donors in the US due to potential transmissible spongiform encephalopathies (TSE) risk [64].
The selection of raw materials (ancillary materials) requires a risk-based approach that assesses potential impact on final cell quality, performance, and characteristics [64]. Key considerations include:
- Quality Grade: Regulators expect the highest quality grade of raw material, with GMP-compliant materials preferred for clinical-facing research
- Animal Origin Free (AOF) Status: Whenever possible, researchers should use AOF reagents that do not contain primary, secondary, or tertiary materials derived from animal sources
- Documentation: Collection of Certificates of Analysis, Certificates of Origin, and supplier regulatory support packages is essential for justifying material choices
Challenges in Neural Differentiation and Regional Patterning
The cerebral cortex consists of a diverse array of cell types including upper and lower layer glutamatergic projection neurons, inhibitory GABAergic interneurons of various fates, and different subtypes of glial cells [62]. Generating all brain-relevant cell types with regional and subtype specificity remains a significant challenge [63]. Recent advances in single-cell analysis of first-trimester human brain tissue have revealed that region-specific transcriptomic signatures first appear in neuroepithelial and radial glia cells at gestational week (GW) 7-8, with regional specification peaking at GW15-18 [66]. This developmental understanding has informed improved differentiation protocols.
Table 1: Key Neural Cell Types and Differentiation Challenges for NDD Modeling
| Neural Cell Type | Developmental Origin | Differentiation Challenge | Relevance to NDDs |
|---|---|---|---|
| Cortical glutamatergic neurons | Dorsal pallium | Achieving specific cortical layer identities (deep vs. upper layers) | SCZ, ASD [63] |
| GABAergic interneurons | Medial/caudal ganglionic eminences | Extended maturation time; subtype specification | SCZ, epilepsy [63] |
| Midbrain dopaminergic neurons | Floor plate precursors | Functional maturation with appropriate markers | ADHD, addiction pathways [63] |
| Hippocampal neurons | Medial pallium | Recapitulating connectivity (e.g., dentate-CA3) | SCZ, memory disorders [63] |
| Astrocytes | Radial glial cells | Functional maturation and diversity | Multiple NDDs [63] |
Automation Strategies for Robust iPSC Bioprocessing
Workforce and Economic Drivers for Automation
Current cell therapy manufacturing relies on numerous ordered steps generally performed by highly qualified operators. A hypothetical 10-day production process involving 300-unit operations running in parallel would require approximately 1,700 full-time employees for a single facility [67]. This workforce is not only challenging to source but economically unsustainable, particularly given high staff attrition rates in the sector—data suggest 70% of employees change jobs every 18 months [67]. Automation directly addresses these challenges by reducing dependence on scarce technical expertise and minimizing training burdens.
Quality and Consistency Enhancements
Automated systems provide significant reductions in product variability, manufacturing errors, and risk of contamination [67]. For iPSC-based NDD models, consistency is particularly crucial as biological variability can confound disease-specific phenotypic observations. A single employee can operate many automated systems simultaneously, increasing the number of tasks completed per unit time and supporting rapid process scalability without the need to train additional staff in manual processing [67]. Furthermore, automated systems typically have a smaller footprint than manual systems, increasing overall facility productivity—a critical consideration for resource-constrained research environments.
Implementation Approaches and Industry Adoption
The transition from manual methods to automated processes depends largely on manufacturers' willingness to share their implementation experiences. As noted by researchers, "Following the open science approach, companies should publish findings regarding the performance of systems under development without compromising intellectual property" [67]. This knowledge sharing is particularly valuable for the research community developing iPSC-based NDD models.
Technology developers must ensure their systems are flexible, as "there is not a unique answer or solution that is suitable for all cell therapies" [67]. Researchers should adopt a risk-based approach to identify which automation strategies would be most beneficial for their specific iPSC modeling applications, particularly when differentiating multiple neural lineages for NDD studies.
Bioprocessing Innovations for Cost-Effective iPSC Manufacturing
Digital Transformation and Data Integration
Digitalization offers transformative potential for bioprocessing, with organizations that have advanced beyond initial testing phases of intelligent automation reporting average cost savings of 32% [68]. The biopharmaceutical sector is increasingly adopting digital technologies, with 75% of executives surveyed reporting routine use of databases for data storage and statistical analysis methods [65]. However, only half apply nonlinear analysis methods such as machine learning (ML) and hybrid models, while digital twins are least established with less than 30% adoption [65].
Centralized process databases represent a key objective in digital transformation, allowing consolidation of all generated data in a standardized format that is consistently updated as new data become available [65]. For iPSC researchers, this approach facilitates better tracking of differentiation efficiency and experimental outcomes across multiple neural differentiations and disease lines.
Table 2: Digital Technology Adoption and Impact in Bioprocessing
| Technology | Current Adoption | Potential Benefits for iPSC Research | Implementation Challenges |
|---|---|---|---|
| Databases & Statistical Analysis | 75% | Structured experimental data management | Data standardization across labs |
| Multivariate Analysis | ~50% | Understanding complex parameter interactions | Requires specialized expertise |
| Machine Learning & Nonlinear Methods | ~50% | Predictive model development for differentiation | Data quality and quantity requirements |
| Digital Twins | <30% | Process optimization without resource-intensive experimentation | Complex model development and validation |
| Online Sensors & Real-time Monitoring | Becoming industry standard | Continuous quality assessment | Integration with existing systems |
Advanced Bioprocessing Methodologies
Several bioprocessing strategies show particular promise for reducing costs in iPSC-based NDD research:
High-Throughput Screening This approach allows rapid testing of numerous variables, such as different media compositions or small molecule concentrations, to identify optimal conditions for iPSC maintenance and neural differentiation. This reduces time spent in trial-and-error phases and accelerates protocol development [69].
Single-Use Systems Traditional stainless-steel bioreactors require extensive cleaning and validation processes that are both time-consuming and costly. Single-use systems eliminate cleaning needs, reduce cross-contamination risks, and offer flexibility for adapting to changing production requirements [69].
Process Analytical Technology (PAT) and Quality by Design (QbD) PAT involves real-time monitoring and control of bioprocesses, enabling more consistent production of iPSCs and their neural derivatives. QbD focuses on designing processes with quality in mind from the outset, reducing the likelihood of costly issues later in development [69].
Supply Chain and Raw Material Management
Strategic management of raw materials presents significant cost-reduction opportunities. Researchers should implement a formal raw material risk assessment once candidate processes are established [64]. This approach identifies and assesses potential risks across a matrix of considerations, resulting in an overall risk score that guides mitigation strategies. Proactive assessment allows for early intervention to mitigate high-risk materials that could compromise research outcomes or regulatory compliance.
Quantitative Cost-Benefit Analysis of Advanced Technologies
The development and manufacturing costs of biologics are significantly higher than those for small molecule drugs—approximately 11 times greater, leading to a daily dose cost approximately 22 times higher [65]. This cost differential directly impacts the feasibility of iPSC-based NDD research programs, particularly for academic institutions and small biotechs.
Intelligent automation delivers cost savings through multiple mechanisms. Organizations report an average cost savings of 32% through reduced labor costs, improved process turnaround times, and decreased errors and rework [68]. These metrics can be quantified specifically for iPSC research:
- Reduced Labor Costs: Calculate human hours freed by automation multiplied by average labor cost per hour
- Improved Process Turnaround: Analyze time saved by automating tasks with corresponding cost reductions
- Reduced Errors and Rework: Quantify costs associated with experimental repetition due to manual errors
Additional financial benefits beyond immediate cost reduction include improved experimental consistency (critical for phenotypic validation in NDD models), enhanced decision-making through automated data analysis, and increased research output through more efficient resource utilization.
Integrated Experimental Protocols for iPSC-Based NDD Research
iPSC Generation and Characterization Protocol
Starting Material Acquisition Begin with somatic cell acquisition from donors with comprehensive documentation. For allogeneic models, complete donor eligibility determination based on medical records review, physical exam, and clinical laboratory testing for relevant communicable diseases [64]. Ensure informed consent covers immortalization, genetic editing, genetic testing, and commercial/research use.
Reprogramming Method Selection Select reprogramming factors and delivery methods considering IP constraints and research objectives. Balance efficiency against safety concerns, with non-integrating methods generally preferred despite potentially lower efficiency [64]. Monitor for genetic and epigenetic abnormalities throughout the process.
Characterization and Banking Implement comprehensive characterization including pluripotency marker assessment, karyotyping, and genetic stability evaluation. Establish master and working cell banks with sufficient documentation for research reproducibility [64].
Neural Differentiation and Regional Patterning
Cortical Glutamatergic Neurons Utilize dual-SMAD inhibition (BMP and TGF-β pathway inhibition) to specify cortical fate from hiPSCs [63]. Subsequent specification into specific cortical layer identities employs timed exposure to patterning factors: deep layer neurons (TBR1+, CTIP2+) appear earlier, while upper layer neurons (BRN2+, CUX1+, SATB2+) emerge later [63] [62]. This temporal patterning mirrors human cortical development in vivo.
GABAergic Interneurons Generate MGE-like neural progenitors using small molecule inhibitors for WNT and SMAD signaling coupled with timed sonic hedgehog (SHH) exposure to induce ventral telencephalic fate [63]. Note that GABAergic interneurons require extended maturation times in vitro, mimicking human neural development timelines [63].
Midbrain Dopaminergic Neurons Direct hiPSCs to differentiate into floor-plate precursors with activation of SHH and WNT/β-CATENIN signaling, followed by differentiation into functional neurons expressing markers TH and PITX3 [63]. These neurons are relevant for studying NDDs with dopaminergic involvement.
Quality Control and Functional Validation
Implement rigorous quality control measures throughout differentiation processes:
- Flow cytometry for cell type-specific markers at multiple timepoints
- Single-cell RNA sequencing to validate population heterogeneity and identity
- Electrophysiological assessment for functional maturation
- Calcium imaging or multielectrode array analysis for network activity
For disease modeling applications, incorporate disease-relevant functional assays that probe the specific neural circuits hypothesized to be affected in the NDD under investigation.
The Scientist's Toolkit: Essential Research Reagents and Platforms
Table 3: Key Research Reagent Solutions for iPSC-Based NDD Modeling
| Reagent Category | Specific Examples | Function in iPSC Neural Differentiation | Considerations |
|---|---|---|---|
| Small Molecule Inhibitors/Activators | Dual-SMAD inhibitors, SHH agonists, WNT modulators | Direct regional patterning of neural progenitors | Concentration and timing critical for specific regional identities |
| Extracellular Matrices | Laminin-521, Synthemax, Vitronectin | Support pluripotency and neural differentiation | Impact on differentiation efficiency and maturation |
| Cell Culture Media | Essential 8, mTeSR, Neural induction media, Regional specification media | Maintain pluripotency or direct differentiation | Lot-to-lot variability; formulation complexity |
| Growth Factors & Cytokines | FGF2, BDNF, GDNF, NT-3 | Support neural progenitor expansion and neuronal maturation | Stability and cost; recombinant human sources preferred |
| Gene Editing Tools | CRISPR/Cas9, TALENs, Base editors | Introduce or correct disease-relevant variants | Off-target effects; delivery efficiency; IP considerations |
Visualizing the Integrated Workflow: From Somatic Cells to Neural Networks
The following diagram illustrates the integrated workflow for generating neural networks from somatic cells, highlighting key decision points and quality control checkpoints in the process:
Diagram 1: Integrated Workflow for iPSC-Based Neural Differentiation
Strategic Implementation Roadmap for Research Laboratories
Implementing comprehensive automation and bioprocessing strategies requires careful planning and phased execution. The following diagram outlines a strategic roadmap for research laboratories:
Diagram 2: Strategic Implementation Roadmap
The field of iPSC-based neurodevelopmental disorder research stands at a pivotal juncture, where technological advances in automation and bioprocessing are enabling unprecedented insights into human-specific disease mechanisms. The convergence of improved differentiation protocols, sophisticated digital monitoring, and intelligent automation creates opportunities to overcome longstanding technical hurdles while managing costs effectively. As single-cell technologies continue to reveal the intricate details of human brain development [66], iPSC-based models will become increasingly refined in their ability to recapitulate disease-relevant phenotypes. By implementing the strategies outlined in this technical guide, researchers can accelerate their investigation of neurodevelopmental disorder mechanisms while optimizing resource utilization, ultimately contributing to improved understanding and therapeutic interventions for these complex conditions.
The integration of human induced pluripotent stem cell (hiPSC) technology with advanced machine learning (ML) frameworks is revolutionizing the modeling of neurodevelopmental disorders (NDDs). This synergy addresses a critical bottleneck in neuroscience: the lack of experimental systems that faithfully recapitulate human neurodevelopment and its perturbations. For decades, progress has been constrained by the limitations of animal models, which, although invaluable, do not capture the intricacies of human-specific developmental processes. The emergence of hiPSCs has opened a new era, enabling the derivation of patient-specific neurons, glia, and three-dimensional (3D) organoid systems that more closely model human physiology and pathology [18]. However, these models generate immense, multi-dimensional datasets that require sophisticated data integration frameworks and machine learning approaches for meaningful biological interpretation. The complexity of these systems is magnified in high-content screening (HCS) platforms, which investigate gene function in an unbiased manner within the cellular context [70]. The ability to multiplex readouts for multi-parametric analysis and utilize cell painting in combination with enhanced machine learning for image analysis has pushed HCS to become an unrivalled technology for discoveries of cell-based phenotypes and gene/protein networks [70]. This technical guide provides a comprehensive framework for implementing machine learning-driven data integration strategies to advance phenotype classification in hiPSC-based NDD research, with particular emphasis on high-content analysis workflows that are transforming our understanding of disease mechanisms.
Machine Learning Frameworks for Biomedical Data Analysis
Selecting appropriate machine learning frameworks is fundamental to building robust predictive models for phenotype classification. These frameworks provide tools, libraries, and interfaces that simplify the process of building, training, testing, and deploying machine learning models, allowing researchers to focus more on problem-solving rather than low-level programming [71]. The field has grown rapidly, with numerous successful technologies leveraging these frameworks for effective development. Several key frameworks have emerged as particularly relevant for handling the complex data structures generated in hiPSC research.
Table 1: Key Machine Learning Frameworks for High-Content Analysis
| Framework | Primary Language | Key Features | Advantages for HCA | Limitations |
|---|---|---|---|---|
| TensorFlow | Python, C++ | Flexible architecture, computational graph visualization, distributed training [71] | Excellent for deploying models across cloud, mobile, and browsers; strong production support [72] | Steep learning curve; frequent updates; limited GPU support [71] |
| PyTorch | Python, C++ | Dynamic computation graphs, native Python integration, eager execution [71] | Greater flexibility using native Python code; fast development cycle; ideal for research prototyping [71] | Lacks model serving in production; limited monitoring interfaces [71] |
| Scikit-learn | Python | Simple API, extensive algorithms for classification, regression, clustering [71] | User-friendly; vast pre-processing abilities; excellent for traditional ML tasks [71] [72] | Lacks native deep learning capabilities; not optimized for very large datasets [71] |
| Hugging Face Transformers | Python | Pre-trained models, consistent API, works with PyTorch/TensorFlow [71] | Revolutionizes NLP tasks in research; simplifies adapting pre-trained models to custom datasets [71] | Focused on transformer architectures; requires significant computational resources [71] |
Framework Selection Considerations
When implementing machine learning for high-content analysis, researchers must consider several factors in framework selection. TensorFlow provides a versatile and strong foundation with a complete library ecosystem, making it suitable for creating classification models, neural networks, regression models, and other machine learning architectures [71]. Its ability to visualize computational graphs offers particular advantages for debugging complex model architectures. However, its steep learning curve can present challenges for beginners. PyTorch, in contrast, offers an intuitive interface that is faster for newcomers to learn, with support for eager execution and greater flexibility through native Python code integration [71]. This makes it particularly valuable for rapid prototyping of new analytical approaches. For traditional machine learning tasks without deep learning requirements, Scikit-learn remains a cornerstone with its user-friendly interface and comprehensive documentation [71]. Its capabilities for data mining and analysis, including support for most supervised learning algorithms and unsupervised learning approaches, make it ideal for preliminary investigations and established analytical pipelines.
Specialized frameworks have also emerged to address specific analytical challenges. Hugging Face Transformers, while technically a library built on top of PyTorch and TensorFlow, has revolutionized Natural Language Processing and is increasingly applied to biological sequence data and scientific text mining [71]. Its extensive collection of pre-trained models and consistent API significantly lowers the barrier to implementing state-of-the-art transformer architectures. When computational resources are limited or research questions do not require complex deep learning architectures, traditional frameworks like Scikit-learn often provide the most efficient path to actionable insights. Ultimately, framework selection should be guided by specific research objectives, available computational resources, and team expertise, with many laboratories adopting a polyglot approach that leverages the strengths of multiple frameworks throughout the analytical workflow.
High-Content Screening and Phenotype Classification in iPSC Models
Experimental Workflow for High-Content Screening
High-content screening (HCS) represents a powerful technological platform for investigating gene function in an unbiased manner within the cellular context [70]. In iPSC-based neurodevelopmental disorder research, HCS enables multiplexed readouts for multi-parametric analysis, which when combined with machine learning, becomes an unrivalled technology for discovering cell-based phenotypes and gene/protein networks [70]. The standard workflow begins with the generation and thorough characterization of iPSC lines to ensure true pluripotency prior to neuronal differentiation and screening experiments [70]. This characterization includes expression of pluripotency markers via immunofluorescence and RT-PCR, Epi-Pluri-Score assessment examining DNA-methylation levels at specific CpG sites, demonstration of spontaneous in vitro differentiation into all three germ layers, maintenance of genomic integrity after reprogramming, and clearance of non-integrating vectors if relevant reprogramming methods were used [70].
Following characterization, iPSCs undergo directed differentiation into relevant neural lineages. For midbrain dopaminergic neurons, this process involves sequential stages with precisely timed addition of specific patterning factors [70]. The differentiation begins with embryoid body formation (Days 0-4) using DMEM/F12 and Neurobasal medium in a 1:1 ratio, supplemented with N2 (1:100) and B-27 (1:50) supplements, L-glutamine (2 mM), penicillin/streptomycin (100 U/ml), and small molecules including Thiazovivin (10 μM, Days 0-2), SB431542 (10 μM), LDN193189 (100 nM), CHIR99021 (0.8 μM), Sonic Hedgehog (100 ng/ml), and Purmorphamine (0.5 μM, from Day 2) [70]. From Days 4-11, neuronal differentiation continues with adjusted concentrations of N2 (1:200) and B-27 (1:100) supplements, with specific factors progressively withdrawn according to established developmental timelines [70]. The resulting day 11 ventral midbrain progenitor cells are then seeded onto 96-well plates pre-coated with poly-L-ornithine, fibronectin, and laminin using automated liquid handling systems to ensure reproducibility [70].
Diagram Title: High-Content Screening Experimental Workflow
Research Reagent Solutions for iPSC-Based Screening
Table 2: Essential Research Reagents for iPSC Neuronal Differentiation and Screening
| Reagent Category | Specific Examples | Function | Application Notes |
|---|---|---|---|
| iPSC Maintenance | mTeSR1/TeSR-E8, Matrigel/Vitronectin XF, ReLeSR [70] | Maintain pluripotency and self-renewal | Quality control critical; batch-to-batch consistency important |
| Neural Induction | SB431542, LDN193189, CHIR99021, Purmorphamine, SHH [70] | Direct differentiation toward neural lineages | Concentrations and timing vary by protocol and target neural subtype |
| Cell Culture Substrates | Poly-L-ornithine, Fibronectin, Laminin [70] | Promote cell attachment and neuronal maturation | Sequential coating often required for optimal neuronal health |
| Staining Reagents | Antibodies against LC3, neuronal markers (TUJ1, MAP2) [70] | Visualize cellular structures and processes | Validation for iPSC-derived neurons essential; multiplexing increases information content |
| Cell Dissociation | TrypLE, Accumax [70] | Gentle dissociation for passaging | Critical for maintaining cell viability while achieving single-cell suspension |
Implementation of high-content screening requires access to specialized instrumentation including an automated liquid handling system for consistent cell seeding and compound administration, an acoustic dispenser for precise compound transfer, and a high-content imaging platform such as the PerkinElmer Opera Phenix system for automated image acquisition [70]. For image analysis, bioinformatics support is recommended to develop robust analytical pipelines. The entire workflow must be optimized for the specific phenotypic endpoints being investigated, with the autophagy marker LC3 serving as one example for investigating autophagosome formation in iPSC-derived neurons [70]. However, the protocol can be adapted to analyze various proteins of interest relevant to specific neurodevelopmental disorders, enabling customized investigation of disease-specific phenotypes.
Computational Approaches for Phenotype Classification
Image Analysis and Feature Extraction
The computational workflow for phenotype classification begins with robust image analysis to extract quantitative features from high-content imaging data. For iPSCs with challenging morphology, segmentation of single-channel phase-contrast images presents particular difficulties. Innovative approaches incorporating CellProfiler-based image analysis pipelines enable segmentation of these challenging samples [73]. When coupled with R-based software solutions to reduce the dimension of time to a single data point, researchers can integrate live imaging data with endpoint analyses while retaining the dynamics of cellular responses [73]. These combined packages allow robust segmentation of iPSCs solely on phase-contrast single-channel images and enable characterization of the response of live-imaged iPSCs to external stimuli, defining cell line-specific phenotypic signatures [73].
Following segmentation, feature extraction generates quantitative descriptors of cellular morphology, texture, and spatial relationships. These features form the basis for subsequent machine learning classification. For iPSC-derived neuronal cultures, relevant features may include neurite outgrowth, branching complexity, soma size, and spatial distribution of specific markers. In the context of neurodevelopmental disorders, studies have identified 663 distinct cellular phenotypes from 71 gene mutations across 31 neurological disorders [74]. These iPS cellular phenotypic traits (iCPTs) can be organized into a taxonomy of nine clusters: decreased cellular processes and products (37%), increased cellular processes and products (35%), impairment of expected cellular functions (5%), increased susceptibility to chemical exposure (5%), presence of abnormal cellular structures (6%), accumulation of molecules (5%), decreased susceptibility to chemical exposure (2%), rescue/recovery from disease phenotypes after chemical treatment (4%), and absence of expected normal phenotypes (1%) [74]. This systematic classification provides a framework for organizing the complex phenotypic landscape observed in NDD models.
Machine Learning Implementation for Phenotype Classification
Diagram Title: Machine Learning Phenotype Classification Pipeline
The implementation of machine learning for phenotype classification involves several methodical steps. Feature selection reduces dimensionality to identify the most informative features, improving model performance and interpretability. Techniques include recursive feature elimination, principal component analysis, and domain knowledge-guided selection. Model training employs various algorithms depending on data characteristics and research questions. Support Vector Machines (SVMs) often perform well for high-dimensional data with clear separation boundaries, while Random Forests provide robust performance with minimal parameter tuning and feature importance metrics. Neural Networks, particularly Convolutional Neural Networks (CNNs), can learn features directly from images, bypassing manual feature engineering [18].
For neurodevelopmental disorder research, these approaches are increasingly combined with multi-omics integration. The challenge is no longer only how to collect data, but how to integrate and interpret it meaningfully across genomics, transcriptomics, proteomics, and metabolomics domains [18]. Computational tools like MetaboLINK, which combines principal component analysis with graphical lasso, exemplify innovations that parse longitudinal metabolomics data to reveal stage-specific metabolic programs during neural differentiation [18]. Similarly, rich functional datasets from electrophysiological recordings, multielectrode array analyses, and calcium imaging can be combined with machine learning to classify subtle phenotypic signatures and accelerate drug screening [18]. This integration of experimental and computational approaches exemplifies the field's shift toward predictive and data-driven frameworks for NDD research.
Advanced Applications and Multi-Omics Integration
Integrating hiPSC Models with Multi-Omics Technologies
The integration of hiPSC-derived models with multi-omics technologies represents the cutting edge of neurodevelopmental disorder research. As experimental systems become more sophisticated, they generate increasingly complex datasets spanning genomics, transcriptomics, proteomics, and metabolomics [18]. The integration of these diverse data types provides unprecedented opportunities to understand the complex pathophysiology of NDDs. For example, in the context of epilepsy research, generating iPSCs from patients harboring novel CLCNKB mutations and differentiating these cells into neurons for transcriptomic profiling has identified differentially expressed genes implicated in epileptogenesis, thereby uncovering gene networks that may contribute to seizure susceptibility [18]. Similarly, studies of Tuberous Sclerosis Complex (TSC) have leveraged hiPSC-derived neurons to interrogate dysregulation of the mTOR signaling pathway, leading to the development of mTORC1-selective inhibitors that rescue hyperexcitability and abnormal neuronal morphology in TSC2−/− neurons [18].
Beyond two-dimensional cultures, three-dimensional brain organoids are increasingly used to capture more complex features of neurodevelopment [18]. These systems recapitulate important aspects of early developmental processes such as progenitor proliferation, neuronal migration, and layer formation, allowing identification of disrupted pathways from cytoskeletal regulation to growth factor signaling that drive cortical malformations [18]. The combination of organoid technology with multi-omics approaches and machine learning creates a powerful platform for identifying previously inaccessible disease mechanisms. However, standardization of differentiation protocols, reproducibility across laboratories, and integration of massive multi-layered datasets continue to be pressing issues that the field must address [18]. Nevertheless, the trajectory is clear: by uniting stem cell biology, multi-omics integration, and computational frameworks, the field is moving toward more predictive, patient-specific, and ultimately actionable models of neurodevelopmental disorders.
Data Integration Challenges and Emerging Solutions
The implementation of data integration frameworks for hiPSC-based research faces several significant challenges. Methodological heterogeneity across studies presents a major obstacle, with analyses revealing striking differences in research practices and reporting standards. Examination of 93 studies of neurological disease models with human iPSCs found that only a small number utilized isogenic lines (considered a rigorous control), only approximately a quarter submitted gene expression profiles to public databases, and just over half used at least more than one control and disease patient line [74]. Notably, only four studies utilized all four of these methodological rigor elements [74]. This heterogeneity complicates cross-study comparisons and meta-analyses.
Emerging solutions to these challenges include the development of specialized data integration platforms. iPhemap (iPS cell phenogenetic map project atlas) provides an open submission, online database to continually catalog disease phenotypes [74]. This platform enables researchers to query and deposit phenotypic information of neurological diseases, creating a growing resource of standardized phenotype-genotype relationships. Similarly, TableGPT2 represents an innovative approach to tabular data integration, featuring a novel table encoder specifically designed to capture schema-level and cell-level information [75]. This model, trained on over 593.8K tables and 2.36M high-quality query-table-output tuples, strengthens the ability to handle ambiguous queries, missing column names, and irregular tables commonly encountered in real-world applications [75]. Such tools are particularly valuable for integrating the complex, structured data generated in high-content screening campaigns.
The future of data integration in iPSC research will likely involve increasingly sophisticated multi-scale modeling approaches that connect molecular phenotypes to cellular and network-level dysfunction. This will require not only technical solutions for data integration but also conceptual frameworks for understanding how perturbations at different biological scales interact to produce disease states. The application of graph neural networks to represent biological knowledge graphs, incorporating genes, proteins, cellular components, and phenotypes, shows particular promise for illuminating these complex relationships. As these technologies mature, they will progressively transform our ability to classify phenotypes, identify disease mechanisms, and ultimately develop targeted interventions for neurodevelopmental disorders.
Human induced pluripotent stem cells (iPSCs) have revolutionized the study of neurodevelopmental disorders (NDDs) by providing patient-specific cellular models that recapitulate the genetic background of complex conditions such as epilepsy, Tuberous Sclerosis Complex, and autism spectrum disorders [18]. However, the tremendous promise of iPSC technology is tempered by significant challenges posed by genetic and epigenetic variability, which can compromise experimental reproducibility and translational validity [76]. This technical guide outlines evidence-based best practices for managing these variability sources through rigorous cell line characterization and banking procedures, specifically framed within the context of NDD research.
The inherent variability in iPSCs manifests across multiple dimensions: genetic and epigenetic variations exist between different iPSC lines, between different passages of the same line, and even between different populations at a specific passage [76]. In NDD research, where subtle phenotypic differences may underlie disease mechanisms, controlling these variabilities becomes paramount. This guide provides a comprehensive framework for establishing quality-controlled iPSC resources that will enhance the reliability of NDD disease modeling, drug screening, and mechanistic studies.
Genetic Variations in iPSCs
iPSC genomes can harbor multiple types of genetic variations that significantly impact their research utility, particularly in neurological disease modeling where precise genotype-phenotype correlations are essential.
Table 1: Types and Origins of Genetic Variations in iPSCs
| Variation Type | Description | Potential Origin | Impact on NDD Research |
|---|---|---|---|
| Aneuploidy | Abnormal chromosome numbers (e.g., trisomy 12, 8, or X) | Culture adaptation; inherited from source cells | Alters dosage of pluripotency genes; may confound disease-specific phenotypes |
| Subchromosomal CNVs | Mb-scale copy number variations | Reprogramming stress; pre-existing in source cells | May affect neurodevelopment genes; potential false positives in disease modeling |
| SNVs | Single nucleotide variations | Reprogramming mutations; low-frequency in source population | Can introduce confounding functional changes in neuronal signaling pathways |
Aneuploidy occurs frequently in cultured pluripotent stem cells, with approximately one in three human iPSC or ESC lines exhibiting karyotype abnormalities [76]. These abnormalities often confer selective growth advantages in vitro—for instance, trisomy 12 increases the dosage of pluripotency genes NANOG and GDF3 [76]. For NDD research, where neuronal differentiation capacity is crucial, such karyotypic abnormalities can profoundly alter developmental trajectories and mask disease-specific phenotypes.
Copy number variations (CNVs) can originate from multiple sources. Pre-existing CNVs in the heterogeneous source cell population can be captured during clonal iPSC derivation, while de novo CNVs may arise from replication stress during reprogramming [76]. Early-passage iPSCs often exhibit CNV mosaicism, which gradually resolves with passaging as certain variations are selected against [76].
Epigenetic Variations in iPSCs
Epigenetic variability represents another significant challenge in iPSC-based disease modeling. The reprogramming process involves extensive epigenetic remodeling, which may be incomplete or variable across different cell lines.
Epigenetic Memory: iPSCs may retain epigenetic signatures of their somatic cell origin, potentially creating lineage-specific differentiation biases [77]. This "epigenetic memory" could facilitate differentiation toward the original lineage while impairing differentiation to other cell types—a critical consideration for NDD research requiring specific neuronal subtypes.
Donor-Specific Patterns: Recent evidence confirms that iPSCs maintain donor-specific epigenetic patterns even after reprogramming, with chromatin accessibility and DNA methylation patterns strongly associated with genetic variation at the iPSC stage [61]. Interestingly, this relationship weakens as cells differentiate, with epigenetic variation increasing substantially in differentiated cell types while becoming less dependent on genetic background [61].
Best Practices for Cell Line Characterization
Identity Authentication and Genetic Characterization
Robust identity authentication is fundamental to ensuring the validity of iPSC-based NDD research.
Short Tandem Repeat (STR) Analysis: STR profiling represents the internationally recognized consensus standard for human cell line authentication [78]. This method provides a genetic fingerprint that can detect cross-contamination between cell lines, a well-documented issue that contributes to erroneous conclusions and publication retractions [78]. The authentication process should compare iPSCs to reference samples from the original donor when possible, or to the earliest passage stocks available [78].
Comprehensive Genetic Analysis: Beyond identity confirmation, detailed genetic characterization is essential for quality assessment in NDD research:
- Karyotyping: Conventional G-banding karyotyping detects gross chromosomal abnormalities but has limited resolution (~5-10 Mb)
- SNP Arrays: Provide genome-wide coverage for detecting CNVs and uniparental disomy
- Whole Genome Sequencing: Offers the most comprehensive assessment of SNVs, indels, and structural variants
Table 2: Genetic and Epigenetic Characterization Methods for iPSCs in NDD Research
| Method | Application | Information Provided | Recommended Frequency |
|---|---|---|---|
| STR Profiling | Identity authentication | Genetic fingerprint; detects cross-contamination | Master Cell Bank (MCB); Working Cell Bank (WCB) |
| Karyotyping | Genomic stability | Chromosomal number and large structural variants | MCB; every 10 passages for ongoing culture |
| SNP Array | CNV detection | Genome-wide copy number variations; genetic ancestry | MCB; when establishing new subclones |
| WGS | Comprehensive variant detection | SNVs, indels, structural variants | MCB for reference-grade characterization |
| ATAC-seq | Epigenetic characterization | Chromatin accessibility landscape | MCB; after critical differentiations |
| DNA Methylation Profiling | Epigenetic stability | Genome-wide methylation patterns | MCB; key differentiation stages |
Functional Characterization for NDD Research
For iPSC lines destined for NDD research, functional characterization of neuronal differentiation capacity is particularly important.
Pluripotency Verification: Standard assessment includes flow cytometry for surface markers (SSEA-4, Tra-1-60, Tra-1-81) and PCR analysis for pluripotency genes (OCT4, NANOG, SOX2) [61].
Neuronal Differentiation Capacity: Given the specific requirements of NDD modeling, specialized differentiation assays should be implemented:
- 2D Neuronal Cultures: Monitor efficiency of cortical neuron differentiation using markers (TUJ1, MAP2, FOXG1)
- 3D Brain Organoids: Assess capacity to form structured neural tissues with appropriate zonation [18]
- Electrophysiological Maturation: Verify functional maturation through multielectrode array or patch-clamp recordings [18]
- Single-Cell RNA Sequencing: Characterize cellular heterogeneity and lineage representation in differentiations
Cell Banking Strategies for iPSCs
Tiered Banking System
A structured, tiered banking approach is fundamental to preserving iPSC line integrity and ensuring long-term experimental reproducibility in NDD research.
Seed Stocks: Immediately following derivation or acquisition, preserve seed vials at the earliest possible passage of stable and consistent cultures [78]. These stocks serve as recovery material in case of initial culture loss and should be minimally expanded to limit accumulation of genetic variations.
Master Cell Bank (MCB): The MCB represents a homogenous collection of vials generated from a single pooled culture, cryopreserved simultaneously to ensure uniformity [78] [79]. This bank serves as the foundational stock for all future work and should undergo comprehensive characterization.
Working Cell Bank (WCB): Derived from one or more vials of the MCB, WCBs provide the immediate source of materials for experimental work [78]. Using WCBs extends the usability of the well-characterized MCB while maintaining consistency across experiments.
Banking Procedures and Documentation
Pooling Strategy: To ensure bank homogeneity, expand cells to sufficient numbers, pool them prior to cryopreservation, and aliquot as a single batch [79]. This approach ensures that characterization data from a single vial is representative of the entire bank.
Documentation and Tracking: Maintain meticulous records for each bank, including passage number, population doublings, cryopreservation date, and reagent lot numbers. This traceability is essential for troubleshooting and understanding experimental variability.
Off-site Storage: Secure a portion of the characterized MCB at an off-site location, preferably in a different geographical region, to guard against loss due to local catastrophic events [78].
Experimental Protocols for Characterization
Protocol: Cell Line Authentication via STR Profiling
Principle: Short Tandem Repeat (STR) analysis examines specific loci with repetitive elements that vary in length between individuals, creating a unique genetic profile [78].
Procedure:
- Extract genomic DNA from iPSCs using a validated method (e.g., column-based purification)
- Amplify 8-16 core STR loci (as specified in ANSI/ATCC ASN-0002-2021 standard) via PCR
- Separate amplification products by capillary electrophoresis
- Analyze fragment sizes to generate an allelic profile
- Compare profile to reference sample (donor tissue or early passage stock)
Interpretation: Match thresholds typically require ≥80% allele congruence for authentication. In NDD research involving multiple lines, regular authentication is crucial to prevent cross-contamination that could compromise long-term studies.
Protocol: Epigenetic Characterization via ATAC-seq
Principle: Assay for Transposase-Accessible Chromatin with sequencing (ATAC-seq) identifies genome-wide regions of open chromatin, providing insights into the regulatory landscape [61].
Procedure:
- Harvest 50,000 viable iPSCs or differentiated neurons
- Prepare nuclei by gentle lysis
- Treat with Tn5 transposase to fragment accessible chromatin regions
- Purify and amplify tagmented DNA
- Sequence using an Illumina platform (minimum 25 million reads/sample)
- Align reads to reference genome and call peaks of accessibility
- Perform differential accessibility analysis between samples
Application in NDD Research: ATAC-seq can reveal disease-specific alterations in chromatin architecture in patient-derived neurons, potentially identifying disrupted regulatory elements in neurodevelopmental disorders [61].
The Scientist's Toolkit: Essential Research Reagents
Table 3: Essential Research Reagents for iPSC Characterization in NDD Research
| Reagent/Category | Specific Examples | Research Application | Considerations for NDD Modeling |
|---|---|---|---|
| Reprogramming Systems | Sendai virus, episomal vectors, mRNA | iPSC generation from patient somatic cells | Non-integrating methods preferred; consider somatic cell type (urinary epithelial cells accessible [61]) |
| Cell Culture Media | mTeSR, StemFlex, E8 | iPSC maintenance | Chemically defined media reduce batch variability; essential for differentiation reproducibility |
| Characterization Antibodies | OCT4, NANOG, SOX2 (pluripotency); TUJ1, MAP2 (neuronal) | Immunocytochemistry, flow cytometry | Validate species cross-reactivity; lot-to-lot consistency critical for quantitative comparisons |
| qPCR Assays | Pluripotency panel; neuronal differentiation markers | Gene expression profiling | Use reference genes stable across differentiation; design assays to avoid SNP interference |
| Differentiation Kits | Commercial neuronal differentiation kits | Standardized neuron generation | Optimize for specific neuronal subtypes relevant to NDD being studied (cortical, GABAergic, etc.) |
| Genomic Analysis Kits | STR profiling kits, WGS library prep | Genetic characterization | Use validated kits meeting standards (ANSI/ATCC ASN-0002) [78] |
Managing Variability in NDD Research Applications
Strategic Approaches for Neurodevelopmental Disease Modeling
The intricate nature of neurodevelopmental processes demands specialized strategies for managing iPSC variability:
Multi-Line Approach: Given the epigenetic variability between iPSC lines from the same donor [61], utilize multiple independent iPSC lines per donor or condition. This approach controls for line-specific variations that might be misinterpreted as disease-relevant phenotypes.
Isogenic Controls: For disease modeling, generate genetically corrected versions of patient-derived iPSCs using CRISPR/Cas9 gene editing [4]. These isogenic pairs represent the ideal control by differing only at the disease-relevant locus while maintaining identical genetic background.
Longitudinal Monitoring: Implement ongoing genetic and epigenetic assessment throughout extended differentiations, particularly for brain organoid cultures that may require 200+ days to mature [4]. Spontaneous genetic changes during prolonged culture could confound disease phenotype interpretation.
Emerging Technologies and Future Directions
Single-Cell Multi-omics: Emerging technologies enabling simultaneous measurement of genetic, epigenetic, and transcriptomic information from single cells will enhance understanding of heterogeneity in iPSC-derived neuronal cultures [18].
Computational Integration: Machine learning approaches applied to multi-omics datasets can identify subtle patterns distinguishing disease-specific variations from background noise [18]. Tools like MetaboLINK help parse longitudinal data to reveal stage-specific metabolic programs during neural differentiation [18].
Village Cultures: Recent innovations like "village editing"—where multiple genetically distinct iPSC lines are cultured together and edited in parallel—enable direct comparison of genetic background effects on disease-relevant mutations [4].
Effective management of genetic and epigenetic variability is not merely a quality control exercise but a fundamental requirement for rigorous iPSC-based modeling of neurodevelopmental disorders. By implementing the comprehensive characterization and banking practices outlined in this guide, researchers can significantly enhance the reliability and reproducibility of their NDD studies. The structured approach to cell line authentication, tiered banking, and ongoing monitoring provides a foundation for generating meaningful data that accurately reflects disease mechanisms rather than technical artifacts. As the field progresses toward clinical applications and more complex disease models, these practices will become increasingly vital for translating iPSC technology into genuine insights into neurodevelopmental disorder pathogenesis and treatment.
Validating iPSC-Based NDD Models: Bridging In Vitro Findings with Clinical Translation and Drug Discovery
The use of human induced pluripotent stem cell (iPSC) models has revolutionized the study of neurodevelopmental disorders (NDDs) by providing unprecedented access to living human neural tissue. However, the value of these models hinges entirely on their biological relevance, making rigorous benchmarking against native human biology an essential component of any credible research program. Establishing robust correlations between in vitro iPSC-derived models and in vivo human biology ensures that observed phenotypes accurately reflect disease mechanisms rather than experimental artifacts. This technical guide provides a comprehensive framework for validating iPSC-based NDD models through multi-modal benchmarking against post-mortem human brain tissue and clinical data, with a specific focus on methodology standardization, quantitative assessment, and clinical translation.
The fundamental challenge in iPSC neuroscience lies in the inherent limitations of in vitro systems, which necessarily lack the full complexity of the developing human brain. While iPSC-derived neurons and brain organoids recapitulate many aspects of human neurodevelopment, the question of whether they authentically model specific NDD pathophysiology requires systematic validation against gold-standard references from actual human tissue and clinical outcomes. This guide outlines standardized approaches for this validation process, incorporating recent advances in single-cell technologies, multi-omics integration, and computational biology to establish quantitative metrics for model fidelity. By implementing these benchmarking strategies, researchers can significantly enhance the predictive validity of their iPSC models for therapeutic development and mechanistic studies of NDDs such as autism spectrum disorder (ASD), intellectual disability, and related neurodevelopmental conditions.
Benchmarking Frameworks and Methodological Approaches
Multi-Omics Integration for Molecular Benchmarking
Integrative analysis of multiple molecular layers provides a powerful framework for benchmarking iPSC models against human post-mortem tissue. High-throughput omics technologies—including transcriptomics, proteomics, epigenomics, and metabolomics—enable comprehensive molecular characterization across experimental systems and native tissue. The statistical challenges inherent in these comparisons are significant, given the high dimensionality, sparsity, batch effects, and complex covariance structures of omics data, which require robust normalization, batch correction, imputation, dimensionality reduction, and multivariate modeling approaches [80].
Several computational frameworks have been developed specifically for cross-system molecular comparisons. The DIABLO (Data Integration Analysis for Biomarker discovery using Latent cOmponents) framework enables integration of multiple omics data types to identify correlated features across platforms, while MOFA (Multi-Omics Factor Analysis) infers a set of common factors driving variation across different data modalities [80]. Similarity network fusion combines multiple omics data types by constructing networks that sample the mutual nearest neighbors, creating a comprehensive view of molecular relationships. For transcriptomic benchmarking specifically, specialized normalization methods including DESeq2's median-of-ratios approach, trimmed mean of M values (TMM) from edgeR, and quantile normalization are essential for mitigating technical variation before meaningful biological comparisons can be made [80].
Table 1: Statistical Methods for Multi-Omics Benchmarking of iPSC Models Against Post-Mortem Tissue
| Method Category | Specific Approaches | Application in Benchmarking | Key Considerations |
|---|---|---|---|
| Normalization Methods | DESeq2 (median-of-ratios), edgeR (TMM), Quantile Normalization | Corrects for technical variation in cross-platform comparisons | Method must be matched to data type; RNA-seq requires different approaches than proteomics |
| Batch Effect Correction | ComBat, Limma's removeBatchEffect(), Mutual Nearest Neighbors (MNN) | Removes systematic technical differences while preserving biological signals | Risk of over-correction; must preserve biologically relevant variation |
| Multivariate Modeling | Penalized regression, Sparse Canonical Correlation Analysis, Partial Least Squares | Identifies coordinated molecular changes across systems | Handles "large p, small n" problem common in omics studies |
| Multi-Omics Integration | DIABLO, MOFA, Similarity Network Fusion | Identifies convergent molecular signatures across omics layers | Reveals pathways disrupted across molecular levels |
Single-Cell and Spatial Transcriptomics for Cellular Validation
Single-cell RNA sequencing (scRNA-seq) and spatially resolved transcriptomics have emerged as particularly powerful tools for benchmarking the cellular composition and spatial organization of iPSC-derived models against human brain development. By decomposing complex tissues into their constituent cell types, these technologies enable precise comparison of the neuronal and glial populations generated in vitro with those present in the developing human brain. The strategic integration of phenotypic information from patients with neurodevelopmental disorders with human cortical snRNA-seq datasets has revealed reproducible cell-specific expression biases for genes associated with specific clinical presentations [81].
For example, analysis of human cortical snRNA-seq datasets spanning from the second trimester of gestation to adulthood has demonstrated enriched expression of genes associated with speech/cognitive delay and seizures in specific cell types, particularly excitatory neurons and microglia [81]. This information illuminates the distinct cortical cell types most likely to be impacted by pathogenic variants, providing critical guidance for benchmarking studies. When evaluating iPSC-derived neural models, researchers should assess whether disease-associated genes show appropriate cell-type-specific expression patterns that mirror those observed in human brain tissue. This approach moves beyond simple presence/absence assessments to evaluate whether the correct cellular context is being established in vitro.
The benchmarking process involves multiple analytical steps, beginning with quality control to remove low-quality cells, followed by normalization to remove technical effects, dimensionality reduction, clustering, and cluster annotation using marker genes established from reference post-mortem datasets [81]. Cell type identities are assigned based on transcriptional similarity to reference cell types from developing human cortex, with particular attention to the emergence of disease-relevant neuronal subtypes and glial populations. Differential abundance testing can then determine whether iPSC models from patients with specific NDDs recapitulate expected alterations in cellular composition observed in post-mortem tissue from affected individuals.
Experimental Protocols for Benchmarking Studies
Protocol 1: Transcriptomic Correlation Analysis Using snRNA-seq
Purpose: To evaluate the transcriptional fidelity of iPSC-derived neural cells by comparison with human post-mortem brain reference atlases.
Materials:
- Human iPSC-derived neural progenitors, neurons, or brain organoids
- Reference snRNA-seq dataset from human post-mortem cortex (comparable developmental stage)
- Single-cell RNA sequencing platform (10X Genomics, Smart-seq2, etc.)
- Computational resources for scRNA-seq analysis (Seurat, Scanpy, etc.)
Methodology:
- Sample Preparation: Differentiate iPSCs toward neural lineage of interest using established protocols. For cortical models, employ dual SMAD inhibition with dorsomorphin (1μM) and SB431542 (10μM) to direct toward forebrain fate, with patterning factors added as needed for specific regional identities [82].
- Single-Cell Sequencing: Dissociate neural cultures or organoids at multiple time points (e.g., day 30, 60, 90, 120) to capture developmental progression. Prepare single-cell suspensions with viability >85%. Process through chosen scRNA-seq platform following manufacturer's protocol, targeting 5,000-10,000 cells per sample.
- Data Processing: Perform quality control to remove cells with <200 genes or >10% mitochondrial reads. Normalize data using SCTransform or similar variance-stabilizing approaches. Integrate with reference post-mortem dataset using mutual nearest neighbor correction or Seurat's anchor-based integration.
- Comparative Analysis: Project iPSC-derived cells into reference-defined cell type space. Quantify similarity using correlation metrics, cluster co-embedding, and cell type classification scores. Assess expression of disease-associated gene sets in corresponding cell types between model and reference.
Validation Metrics:
- Transcriptomic similarity score (correlation coefficient with reference cell types)
- Percentage of cells correctly classifying to expected neural lineages
- Expression concordance of cell type-specific marker genes
- Developmental trajectory alignment with reference maturation patterns
Protocol 2: Functional Benchmarking Using Electrophysiological and Synaptic Assessment
Purpose: To evaluate the functional maturation and synaptic properties of iPSC-derived neurons against known physiological benchmarks from human neurons.
Materials:
- iPSC-derived neurons (≥60 days differentiation)
- Multi-electrode array (MEA) system or patch-clamp rig
- Immunocytochemistry reagents for synaptic markers
- Human fetal neural tissue-derived neural stem/progenitor cells (hN-NSPCs) as reference standard [82]
Methodology:
- Neuronal Differentiation: Generate cortical neurons using established protocols with neurosphere intermediate stage. Culture for extended durations (≥90 days) to promote maturation, with medium changes every 3-4 days using Neurobasal Plus Medium containing B27 Plus Supplement, GlutaMAX, and brain-derived neurotrophic factor (BDNF, 20ng/mL) [82].
- Synaptic Marker Analysis: At multiple time points, fix cells and immunostain for presynaptic (SYN1, VGLUT1) and postsynaptic (PSD-95, drebrin) markers. Quantify puncta density, size, and colocalization. Compare maturation timeline to reference data from human brain tissue.
- Functional Assessment: Plate neurons on MEA chips or coverslips for patch-clamp recording. For MEA, record spontaneous activity weekly from day 30 onward. For patch-clamp, assess passive membrane properties, action potential characteristics, and synaptic currents.
- Data Analysis: Compare functional properties to published values from human neurons where available. For cortical neurons, evaluate emergence of network synchronization and response to pharmacological agents (GABAergic and glutamatergic receptor antagonists).
Validation Metrics:
- Resting membrane potential (target: -50 to -60 mV)
- Action potential amplitude (target: ≥80 mV)
- Presence of spontaneous postsynaptic currents
- Network bursting activity in MEA (after ≥70 days)
- PSD-95 puncta density and colocalization with presynaptic markers
Table 2: Key Maturation Markers for Benchmarking Neuronal Development in iPSC Models
| Maturation Stage | Structural Markers | Functional Assays | Expected Timeline in iPSC-Derived Neurons |
|---|---|---|---|
| Early Neurogenesis | SOX2, PAX6, NESTIN | Not applicable | Days 10-30 |
| Neuronal Commitment | TUJ1, MAP2, DCX | Not applicable | Days 20-40 |
| Synaptic Formation | SYN1, VGLUT1, PSD-95 | Miniature postsynaptic currents | Days 40-80 |
| Network Maturation | PSD-95, drebrin A, GAD67 | Synchronized network bursting | Days 70+ |
Pathway and Workflow Visualization
Experimental Benchmarking Workflow
Synaptic Maturation Pathway for Functional Benchmarking
The Scientist's Toolkit: Essential Research Reagents and Platforms
Table 3: Essential Research Reagents and Platforms for Benchmarking Studies
| Reagent/Platform | Specific Examples | Function in Benchmarking | Technical Considerations |
|---|---|---|---|
| scRNA-seq Platforms | 10X Genomics, Smart-seq2 | Cellular resolution transcriptomic profiling | 10X for throughput, Smart-seq2 for sensitivity |
| Spatial Transcriptomics | 10X Visium, MERFISH | Tissue architecture preservation | Bridges cellular and tissue-level validation |
| Reference Datasets | BrainSpan, PsychENCODE, Human Cell Atlas | Developmental transcriptome benchmarks | Must match developmental stage |
| Multi-omics Integration Tools | DIABLO, MOFA+, Seurat V5 | Cross-platform data integration | Handles technical batch effects |
| Electrophysiology Systems | Multi-electrode arrays, Patch-clamp | Functional neuronal assessment | Extended maturation required for network activity |
| Synaptic Markers | PSD-95, VGLUT1, SYN1, Gephyrin | Structural synapse validation | Postsynaptic markers lag in iPSC neurons |
| Cell Surface Antigens | CD11B, GFAP, NCAM | Purity assessment and contamination check | Flow cytometry or immunocytochemistry |
Clinical Correlation and Therapeutic Validation
The ultimate validation of any disease model lies in its ability to predict clinical outcomes and response to therapeutic interventions. For iPSC models of NDDs, this involves establishing quantitative relationships between in vitro phenotypes and donor clinical characteristics. In a landmark study of sporadic amyotrophic lateral sclerosis (ALS), researchers established a direct correlation between the rate of neurite degeneration in iPSC-derived motor neurons and donor survival time, providing critical face validity for the model [83]. This approach can be adapted for NDD research by correlating specific molecular or functional phenotypes with clinical severity scores, age of onset, or treatment response.
Therapeutic validation represents another powerful benchmarking approach. Large-scale drug screening in iPSC-derived motor neurons from 100 sporadic ALS patients demonstrated that 97% of drugs previously tested in ALS clinical trials failed to mitigate neurodegeneration in the patient-derived models, accurately reflecting clinical trial outcomes [83]. This striking concordance validates the predictive capacity of properly benchmarked iPSC models. For NDD research, this approach involves testing medications with known clinical efficacy (e.g., risperidone for ASD-associated irritability) to determine whether patient-derived models show corresponding responses.
To establish robust clinical correlations, researchers should implement standardized clinical data collection including detailed phenotypic characterization, standardized severity metrics (e.g., ADOS for ASD, Vineland Adaptive Behavior Scales), and longitudinal outcome measures. These clinical data points can then be correlated with quantitative in vitro phenotypes such as synaptic density, neuronal activity patterns, transcriptomic signatures, or pharmacological responses. The integration of rich genotype-phenotype data from large cohorts like the Deciphering Developmental Disorders study with iPSC phenotyping provides unprecedented opportunities to identify cellular phenotypes associated with specific genetic variants and clinical presentations [81].
Benchmarking iPSC-based models of neurodevelopmental disorders against human post-mortem tissue and clinical data is not merely a quality control measure but a fundamental requirement for generating biologically meaningful insights. The frameworks and methodologies outlined in this guide provide a roadmap for establishing rigorous correlations between in vitro observations and human biology, thereby enhancing the validity and predictive power of iPSC-based NDD research. As the field advances, several emerging technologies promise to further strengthen these benchmarking approaches.
Spatially resolved transcriptomics and proteomics will enable more precise architectural comparisons between iPSC-derived organoids and the complex tissue organization of the developing human brain. Advanced computational methods, including deep learning approaches for data integration and pattern recognition, will enhance our ability to identify subtle but biologically significant correspondences between models and native tissue. Additionally, the generation of increasingly comprehensive reference atlases of human brain development across diverse populations will provide more nuanced benchmarks for evaluating model fidelity. By consistently implementing these benchmarking strategies, the research community can ensure that iPSC-based models fulfill their potential to unravel the complex mechanisms of neurodevelopmental disorders and accelerate the development of effective therapeutics.
The high failure rate of drug candidates in clinical trials, particularly for central nervous system (CNS) disorders, represents a significant challenge in biomedical research [84]. This failure often stems from the poor predictive validity of traditional animal and immortalized cell line models, which do not fully recapitulate human physiology, genetic diversity, or disease etiology [84] [85]. The emergence of human induced pluripotent stem cell (iPSC) technology has revolutionized this landscape by providing a robust platform for generating patient-specific human cells in vitro.
Induced pluripotent stem cells (iPSCs) are derived from reprogrammed adult somatic cells, such as fibroblasts or blood cells, through the induced expression of reprogramming transcription factors like OCT3/4, SOX2, KLF4, and c-MYC [85] [86]. These cells can theoretically differentiate into any cell type in the human body, offering an unprecedented opportunity to create genetically accurate human models for disease study and drug development [84] [85]. For neurodevelopmental disorders (NDDs), where access to functional human neural tissue is severely limited, iPSC-derived models provide particularly valuable insights into disease mechanisms and therapeutic responses [20].
This technical guide examines the predictive validity of iPSC models in high-throughput drug screening and toxicity assessment, focusing specifically on applications within neurodevelopmental disorder research. We cover the fundamental workflows, key experimental protocols, quantitative performance data, and essential research tools that establish iPSC technology as a transformative approach in preclinical drug discovery.
iPSC Technology Workflow for Neurodevelopmental Research
The general workflow for utilizing iPSCs in modeling neurodevelopmental disorders involves several critical stages, from patient recruitment to data analysis. The process fundamentally introduces the "human context" early in the drug discovery pipeline, enabling researchers to study disease mechanisms and test therapeutic interventions in a physiologically relevant human cellular environment [84].
Figure 1: Comprehensive workflow for using iPSC models in neurodevelopmental disorder research and drug screening
Patient-Derived iPSC Generation and Neural Differentiation
The process begins with collecting somatic cells from patients with specific neurodevelopmental disorders and healthy controls. These cells are reprogrammed into iPSCs using defined factors, typically through non-integrating methods such as Sendai virus or mRNA transfection to minimize genomic alterations [84] [86]. The resulting iPSC lines carry the complete genetic background of the donor, including any disease-relevant mutations [85] [20].
For neurodevelopmental disorder research, iPSCs are differentiated into neural lineages using established protocols. Common approaches include:
- Dual-SMAD inhibition: Using small molecule inhibitors of BMP and TGFβ signaling to selectively enrich for neural ectoderm lineage progenitors [84].
- Neural progenitor cell (NPC) expansion: Generating expandable, self-renewable multipotent NPCs as an intermediate population that can subsequently differentiate into astrocytes, oligodendrocytes, and functional neurons [84].
- 3D organoid formation: Allowing self-organization into brain region-specific organoids that better recapitulate the cellular diversity and organization of the developing brain [85] [20].
These differentiation strategies can be tailored to generate specific neural subtypes and circuitries most relevant to the neurodevelopmental disorder being studied, such as cortical neurons for autism spectrum disorders or dopaminergic neurons for certain forms of intellectual disability [20].
High-Throughput Screening Methodologies
High-throughput screening (HTS) using iPSC-derived models requires specialized approaches to handle the complexity of neural cells while maintaining scalability and reproducibility. Several technological advances have enabled more effective screening platforms.
Phenotypic Screening Approaches
Cell Painting is an image-based assay that uses multiple fluorescent dyes to label various cellular components, allowing for the quantification of hundreds of morphological features in response to compound treatment [87]. This approach has been successfully adapted for iPSC screening, where it can detect variable drug responses across donor-specific lines. For example, a recent study demonstrated that a cohort of 28 hiPSC lines from different donors could be screened in their pluripotent state using high-throughput Cell Painting assays, with variable phenotypic responses detected across a wide range of clinically approved drugs [87].
High-content analysis of iPSC-derived neural models provides multidimensional data on disease-relevant phenotypes, including neurite outgrowth, synaptic density, neuronal network activity, and glial cell functions. These analyses can be performed in both 2D cultures and 3D organoid systems, with the latter offering the advantage of capturing more complex cellular interactions [84] [88].
Functional and Molecular Screening
Calcium imaging and multi-electrode arrays (MEAs) are used to assess neuronal functional activity and network formation, providing crucial functional readouts for neurodevelopmental disorders [20]. These approaches can detect subtle alterations in neural circuitry and synaptic function in patient-derived models and assess how therapeutic compounds restore normal activity patterns.
Automated myelination quantification has been developed for 3D BrainSphere models to screen for developmental neurotoxicity. This approach uses flow cytometry and high-throughput imaging to objectively quantify myelin disruption after exposure to potential toxicants, addressing a critical gap in developmental neurotoxicity testing [89].
Proteomic and transcriptomic profiling integrated with screening data provides mechanistic insights into drug responses. For example, quantitative mass spectrometry-based proteomics can identify protein expression changes underlying the variable phenotypic responses observed in Cell Painting assays [87].
Experimental Protocols for Key Applications
Protocol 1: High-Throughput Compound Screening Using iPSC-Derived Neural Progenitors
This protocol outlines a standardized approach for screening compound libraries using iPSC-derived neural progenitor cells (NPCs) in 384-well format, adapted from established methodologies [84] [87].
Materials:
- iPSC-derived NPCs (patient-specific and isogenic controls)
- 384-well tissue culture plates
- Neural expansion medium
- Compound library dissolved in DMSO
- Cell Painting dyes (Mitotracker, Phalloidin, Hoechst, etc.)
- High-content imaging system
- Data analysis software (e.g., CellProfiler, R packages)
Procedure:
- Expand NPCs in neural expansion medium supplemented with EGF and FGF2 until 70-80% confluent.
- Harvest NPCs using gentle enzymatic dissociation and seed at 5,000 cells/well in 384-well plates.
- Allow cells to adhere for 24 hours in a humidified 37°C, 5% CO2 incubator.
- Treat with compound library using automated liquid handling systems, including DMSO vehicle controls and reference compounds.
- After 72 hours of treatment, fix cells with 4% paraformaldehyde for 15 minutes at room temperature.
- Permeabilize with 0.1% Triton X-100 for 10 minutes.
- Stain with Cell Painting dye cocktail according to established protocols [87].
- Image plates using a high-content imaging system with 20x objective.
- Extract morphological features using image analysis software.
- Normalize data and perform statistical analysis to identify hit compounds.
Validation: Include reference compounds with known effects on neural development in each screening batch to assess assay performance. Calculate Z-factor to confirm robustness.
Protocol 2: Developmental Neurotoxicity Screening Using 3D Myelination Models
This protocol describes a specialized approach for assessing compound effects on myelination, a critical process in neurodevelopment, using 3D human BrainSphere models [89].
Materials:
- iPSC-derived neural lineages in 3D culture
- Myelination medium
- Known myelin disruptors (e.g., cuprizone) for positive controls
- Negative controls (e.g., ibuprofen, acetaminophen)
- Anti-myelin basic protein (MBP) antibody
- Flow cytometry system or high-content imager
- Analysis software
Procedure:
- Generate iPSC-derived BrainSpheres using established 3D differentiation protocols.
- Maintain BrainSpheres in myelination-promoting medium for 60-90 days to allow compact myelin formation.
- Treat BrainSpheres with test compounds, positive controls, and negative controls for 14 days.
- Dissociate BrainSpheres to single-cell suspension using enzymatic digestion.
- Fix cells and stain with anti-MBP antibody and appropriate secondary antibody.
- Analyze myelin content using flow cytometry or high-content imaging.
- Quantify the percentage of MBP-positive cells and fluorescence intensity.
- Normalize data to vehicle controls and calculate statistical significance.
Validation: The assay should demonstrate robust disruption by positive controls (cuprizone) while showing no significant effects from negative controls, establishing specificity and sensitivity [89].
Quantitative Assessment of Predictive Validity
The predictive validity of iPSC models can be evaluated through their performance in identifying known toxic compounds and predicting clinical outcomes. The following tables summarize key quantitative data on the predictive capabilities of various iPSC-based testing platforms.
Table 1: Predictive Performance of Stem Cell-Based Developmental Toxicity Assays
| Assay Type | Species | Accuracy | Sensitivity | Specificity | References |
|---|---|---|---|---|---|
| Mouse Embryonic Stem Cell Test (mEST) | Mouse | 78% | NA | NA | [90] |
| Rat Micromass Test | Rat | 70% | NA | NA | [90] |
| Rat Whole-Embryo Culture | Rat | 80% | NA | NA | [90] |
| Zebrafish Embryotoxicity Test | Zebrafish | 72% | NA | NA | [90] |
| FACS-EST | Mouse | 100% | NA | NA | [90] |
| Molecular-EST | Mouse | 72% | 76% | 69% | [90] |
| High-Throughput Imaging (hESC) | Human | 94% | 97% | 92% | [90] |
| Untargeted Metabolomics | Human | 88% | 80% | 100% | [90] |
| 3D Myelination Model | Human | Robust quantification demonstrated | Specific for myelin disruptors | No effects from negative controls | [89] |
Table 2: Applications of iPSC-Derived Models in Neurodevelopmental Disorder Research
| Disorder | Gene Mutation | Key Phenotypes in iPSC Models | Drug Screening Applications | References |
|---|---|---|---|---|
| Fragile X Syndrome | FMR1 CGG repeats (>200) | Hypermethylation of FMR1, defective neurite outgrowth, aberrant differentiation | Demethylating agents, mGluR antagonists | [20] |
| Rett Syndrome | MECP2 mutations | Altered neuronal maturation, decreased synaptic density, reduced network activity | IGF-1, gentamicin readthrough compounds | [20] |
| Dravet Syndrome | SCN1A mutations | Altered sodium current, hyperexcitable neuronal networks | Cannabidiol, serotonin modulators | [20] |
| Phelan-McDermid Syndrome | SHANK3 deletions | Impaired synaptic function, reduced neuronal activity | IGF-1, growth factors, TRKB agonists | [20] |
| Alzheimer's Disease | PSEN1, PSEN2, APP | Amyloid-β production, tau pathology, synaptic dysfunction | BACE inhibitors, gamma-secretase modulators | [84] [85] |
| Parkinson's Disease | SNCA, LRRK2, GBA | Increased α-synuclein, mitochondrial dysfunction, dopaminergic neuron loss | Glucocerebrosidase enhancers, LRRK2 inhibitors | [85] |
Signaling Pathways in Neurodevelopmental Disorders
Understanding the signaling pathways disrupted in neurodevelopmental disorders is essential for developing targeted screening approaches. The following diagram illustrates key pathways that can be modeled in iPSC-derived neural systems.
Figure 2: Key signaling pathways disrupted in neurodevelopmental disorders and modeled in iPSC systems
Essential Research Reagents and Tools
The successful implementation of iPSC-based screening requires specialized reagents and tools. The following table outlines key resources for establishing these platforms.
Table 3: Essential Research Reagents for iPSC-Based Screening Platforms
| Reagent Category | Specific Examples | Function | Commercial Sources |
|---|---|---|---|
| Reprogramming Systems | Sendai virus vectors, mRNA kits, episomal plasmids | Somatic cell reprogramming to iPSCs | Thermo Fisher, Takara Bio, Stemcell Technologies |
| Neural Differentiation Kits | Dual-SMAD inhibitors, neural induction media | Directed differentiation to neural lineages | Stemcell Technologies, Thermo Fisher, Takara Bio |
| Cell Culture Matrices | Geltrex, Matrigel, Laminin-521 | Extracellular matrix for iPSC and neural culture | Corning, Thermo Fisher |
| Characterization Antibodies | Anti-OCT4, SOX2, NANOG (pluripotency); PAX6, NESTIN, TUJ1 (neural) | Validation of pluripotency and neural differentiation | MilliporeSigma, Abcam, BD Biosciences |
| Functional Assay Kits | Calcium imaging dyes, multi-electrode arrays, ATP assays | Assessment of neuronal function and viability | Thermo Fisher, Axion Biosystems, Promega |
| Cell Painting Dyes | Mitotracker, Phalloidin, Hoechst, WGA, Concanavalin A | Multiplexed morphological profiling | Thermo Fisher, MilliporeSigma |
| 3D Culture Systems | Ultra-low attachment plates, spinning bioreactors | Support for organoid and spheroid culture | Corning, Stemcell Technologies |
| Commercial iPSC-Derived Neural Cells | iCell Neurons, iCell Astrocytes, BrainSpheres | Ready-to-use neural cells for screening | Fujifilm CDI, Axol Bioscience, Ncardia |
Challenges and Future Directions
Despite significant advances, several challenges remain in optimizing the predictive validity of iPSC models for high-throughput screening. Addressing these limitations will be crucial for further enhancing the translational value of this technology.
Technical Challenges include the optimization of large-scale, cost-effective culture of iPSC-derived neural cells; the standardization of differentiation protocols across different genetic backgrounds; and the improvement of reproducibility in phenotypic assays [84]. Incorporation of aging-related phenotypes into relatively immature neuronal models also represents a significant hurdle for modeling late-onset aspects of neurodevelopmental disorders [84].
Biological Complexities such as the limited representation of blood-brain barrier function, incomplete maturation of neural circuits in vitro, and absence of systemic immune interactions in current models reduce their physiological relevance [90]. Multi-lineage approaches that incorporate microglia and vascular cells may address some of these limitations [88].
Future advancements will likely focus on the development of more complex multi-lineage models using organ-on-chip technologies and humanized animal systems [88]. Integrated efforts to assemble patient iPSC-derived models with deeply annotated clinicopathological data, along with molecular and drug-response signatures, may aid in patient stratification and clinical trial success [84]. Continued advancement is expected to help fully 'humanize' the process of CNS drug discovery and toxicity assessment.
In conclusion, iPSC-based models demonstrate increasing predictive validity for drug screening and toxicity assessment in neurodevelopmental disorder research. The integration of patient-specific genetic backgrounds with increasingly sophisticated neural differentiation protocols and high-throughput screening technologies positions this approach as a transformative tool in the preclinical drug development pipeline.
The FDA Modernization Act 2.0, signed into law in December 2022, marks a pivotal shift in the U.S. regulatory landscape for drug development [91]. This legislation, driven by recognition of the high failure rate of therapeutics validated in animal models, explicitly removes the long-standing mandate for animal testing and encourages the use of New Approach Methodologies (NAMs) for safety and efficacy testing [91] [92]. For researchers focused on induced pluripotent stem cell (iPSC) modeling of neurodevelopmental disorders (NDDs), this creates an unprecedented opportunity. The Act provides a regulatory pathway to employ patient-specific iPSC-derived models, which more accurately recapitulate human-specific brain development and pathophysiology, thereby accelerating the translation of basic research into clinically relevant therapies [93] [94] [95]. This guide details the core provisions of the Act, the NAMs most relevant to iPSC-NDD research, and practical experimental protocols to navigate this new paradigm successfully.
Decoding the FDA Modernization Act 2.0 and Subsequent Policy Shifts
The FDA Modernization Act 2.0 fundamentally revised the Federal Food, Drug, and Cosmetic Act of 1938, eliminating the requirement for animal testing for investigational new drug (IND) applications [91] [96]. Instead, the law now permits sponsors to use "certain alternatives," including cell-based assays, organoids, microphysiological systems (MPS), and computer models [91] [92]. This legislative change paved the way for a significant policy announcement from the FDA in April 2025, which outlined a ambitious roadmap to actively phase out animal testing requirements [97] [98] [99].
Key Regulatory Milestones and Definitions
Table: Key Definitions in the New Regulatory Landscape
| Term | Definition | Relevance to iPSC-NDD Research |
|---|---|---|
| New Approach Methodologies (NAMs) | A broad category of innovative, human-biology-based testing methods. Includes in vitro (cell-based), in silico (computer model), and ex vivo systems [97] [99]. | The overarching framework for using iPSC-derived models in regulatory submissions. |
| Microphysiological Systems (MPS) | Engineered in vitro models that mimic the structure and function of human tissues and organs. Often referred to as "organ-on-a-chip" systems [91] [92]. | Enables creation of complex, multi-cellular neural models with physiologically relevant cues. |
| In Silico Models | Computer simulations and AI-based models used to predict drug toxicity, metabolism, and biological activity [97] [91]. | Can be used to analyze sequencing data from iPSC models and predict patient-specific drug responses. |
| Human Induced Pluripotent Stem Cells (iPSCs) | Pluripotent stem cells derived from reprogrammed somatic cells (e.g., skin fibroblasts, blood cells) [93] [95] [3]. | The foundational biological material for creating patient- and disease-specific NAMs. |
The FDA's 2025 announcement emphasizes an immediate focus on monoclonal antibodies and other biologics, with a plan to expand to other drug classes [97] [98]. The agency is incentivizing this transition by offering faster approval times and streamlined reviews for sponsors who integrate robust NAM data into their applications [96]. Furthermore, the FDA will now consider preexisting, real-world safety data from countries with comparable regulatory standards where a drug has already been studied in humans, reducing redundant testing [97] [99].
Core NAMs for iPSC-Based Neurodevelopmental Disorder Research
The mechanistic study of NDDs requires models that can recapitulate the early, dynamic processes of human brain development. Patient-derived iPSCs provide a unique platform for this, as they retain the patient's complete genetic signature and can be differentiated into the relevant neuronal and glial cell types affected in these disorders [93] [94]. The following NAMs are particularly powerful for this field.
Key iPSC-Derived Model Systems
Table: Essential iPSC-Derived Models for NDD Research
| Model System | Description | Application in NDD Research |
|---|---|---|
| 2D Neural Cultures | Monolayers of a specific neuronal subtype (e.g., cortical, dopaminergic) or glial cells derived from iPSCs [93] [95]. | Ideal for high-content imaging, electrophysiology (patch-clamp), and high-throughput drug screening on a single-cell type. |
| 3D Brain Organoids | Self-organizing, three-dimensional structures that contain multiple neural cell types and exhibit primitive tissue-like architecture [93] [3]. | Models complex cell-cell interactions, neural network formation, migration defects, and the effects of tissue organization. |
| Organ-on-a-Chip (OoC) | Microfluidic devices that culture iPSC-derived neural cells in a controlled environment, often allowing for vascular perfusion and integration with other organ models [91]. | Studies the role of fluid flow, shear stress, and blood-brain barrier function in NDDs; enables modeling of multi-organ interactions. |
The Scientist's Toolkit: Key Reagents for iPSC-NDD Research
Table: Essential Research Reagents for iPSC Modeling of NDDs
| Item | Function in Experimental Protocol |
|---|---|
| Yamanaka Factor Reprogramming Kit | Commercial kits containing vectors (often non-integrating Sendai virus or plasmids) for the expression of OCT4, SOX2, KLF4, and c-MYC to generate iPSCs from patient somatic cells [95] [3]. |
| Neural Induction Medium | A defined cocktail of small molecules and growth factors (e.g., SMAD inhibitors, FGF2) to direct the differentiation of iPSCs into neural progenitor cells (NPCs) [93] [95]. |
| Neuronal Maturation Supplements | Components such as BDNF, GDNF, NT-3, and cAMP added to culture media to promote synaptic maturation, neurite outgrowth, and functional network activity in derived neurons [93]. |
| CRISPR/Cas9 System | Genome engineering tools to create isogenic control lines (correcting a patient mutation) or introduce disease-associated mutations into healthy iPSCs, crucial for confirming genotype-phenotype relationships [94]. |
| Cell Village Barcoding Kit | Pools of genetic barcodes used to label iPSCs from multiple donors, allowing them to be cultured together in a single "village" to minimize batch effects in large-scale drug screening [91]. |
Experimental Framework: Implementing NAMs for NDD Mechanistic Studies
This section outlines a core experimental workflow for utilizing iPSC-NAMs to investigate the pathophysiology of NDDs, aligned with the regulatory and scientific principles of the FDA Modernization Act 2.0.
Workflow Diagram: From Patient Cell to Therapeutic Insight
Detailed Experimental Protocols
Protocol 1: Generation and Validation of Patient-Derived iPSCs
- Reprogramming Method Selection: Use a non-integrating method, such as Sendai virus or episomal plasmids, to deliver the reprogramming factors (OCT4, SOX2, KLF4, c-MYC) to patient somatic cells (e.g., skin fibroblasts or peripheral blood mononuclear cells) [95]. This avoids genomic integration and potential insertional mutagenesis.
- Culture Conditions: Maintain emerging iPSC colonies on feeder cells or in feeder-free conditions using defined matrices. Culture in essential pluripotency-supporting media such as mTeSR or StemFlex.
- Validation Steps:
- Pluripotency Confirmation: Immunocytochemistry for key pluripotency markers (OCT4, SOX2, NANOG) and demonstration of differentiation potential into all three germ layers (ectoderm, mesoderm, endoderm) [3].
- Genetic Integrity: Perform karyotype analysis (G-banding) and whole-genome sequencing to ensure no major chromosomal abnormalities or mutations were acquired during reprogramming.
- Line Authentication: Short tandem repeat (STR) profiling to confirm cell line identity matches the original donor.
Protocol 2: Differentiation into 2D Neural Cultures and 3D Brain Organoids
2D Neural Culture:
- Neural Induction: Direct iPSCs toward a neural fate using dual SMAD inhibition (e.g., LDN-193189 and SB431542) for approximately 10-14 days to generate neural progenitor cells (NPCs) [93] [95].
- Neuronal Differentiation: Plate NPCs and switch to a neuronal differentiation medium containing BDNF, NT-3, and ascorbic acid to promote terminal differentiation into glutamatergic, GABAergic, or other specific neuronal subtypes over 4-8 weeks [93].
- Characterization: Analyze by immunostaining for neuronal markers (TUJ1, MAP2), synaptic markers (PSD95, Synapsin), and subtype-specific markers (vGLUT1, GAD67).
3D Brain Organoid Generation (Embryoid Body Method):
- EB Formation: Dissociate iPSCs into single cells and aggregate them in low-adhesion U-bottom plates to form embryoid bodies (EBs) in media containing BMP and TGF-β inhibitors [93].
- Neural Induction: Transfer EBs to a neural induction medium to promote neuroectodermal fate. Embed EBs in Matrigel droplets to provide a 3D scaffold for structured growth.
- Maturation: Place Matrigel-embedded organoids in a spinning bioreactor or orbital shaker for long-term culture (months) to improve nutrient exchange and promote complex cytoarchitecture, including cortical layer formation [93].
Protocol 3: Phenotypic Analysis for NDD Pathophysiology
- Transcriptomic/Epigenomic Profiling: Use single-cell RNA sequencing (scRNA-seq) and Assay for Transposase-Accessible Chromatin (ATAC-seq) on differentiating neurons or organoids to identify disease-specific gene expression patterns and chromatin accessibility changes at multiple developmental time points [93] [91].
- Functional Electrophysiology: Perform multi-electrode array (MEA) recordings on 2D cultures or sliced organoids to assess network-level activity, including synchronicity, burst patterns, and network complexity, which are often aberrant in NDDs [93] [94].
- High-Content Image Analysis: Use automated microscopy and analysis software to quantify neuronal morphology (neurite length, branching, soma size) and synaptic density in high-throughput formats for drug screening [94].
Data Integration and Regulatory Strategy
The final, crucial step is to integrate data from these iPSC-NAMs with other acceptable methods to build a compelling case for an IND application.
- In Silico Validation: Use AI and machine learning models to predict the toxicity and metabolism of candidate compounds identified in your screens, based on their chemical structure and your experimental data [91] [96].
- Leverage Real-World Data (RWD): For repurposed drugs or drugs with existing human data from other countries, incorporate this clinical information to support safety and efficacy claims, as explicitly endorsed by the FDA's new policy [97] [99].
- Pilot Programs: Engage with the FDA early through its pilot programs for specific drug classes, such as monoclonal antibodies, to gain experience and feedback on your NAM-based data package under the new regulatory framework [97] [96].
The confluence of the FDA Modernization Act 2.0 and advanced iPSC-based NAMs has created a transformative moment for research into neurodevelopmental disorders. By adopting the detailed experimental frameworks and regulatory strategies outlined in this guide, scientists can move beyond the limitations of animal models and directly interrogate the human-specific pathophysiology of NDDs. This paradigm shift promises to de-risk drug development, accelerate the discovery of meaningful therapies, and ultimately fulfill the great expectations held for iPSC technology in alleviating human disease.
The advent of induced pluripotent stem cells (iPSCs) has revolutionized preclinical drug development by providing unprecedented access to human-specific neural cells for disease modeling and compound screening. By capturing the complete genetic background of patients, iPSC-derived neural cells address a critical translational gap in neuroscience drug discovery, where over 90% of candidates for central nervous system (CNS) disorders fail clinical trials [100]. This whitepaper examines key case studies demonstrating the practical application of iPSC-derived neural models in pharmaceutical development, with particular focus on their utility for elucidating neurodevelopmental disorder mechanisms. These human-relevant systems enable researchers to move beyond animal models that often poorly recapitulate human disease pathophysiology and immortalized cell lines that lack phenotypic fidelity [101] [102]. The case studies that follow highlight how iPSC technology is being deployed across the drug development pipeline, from initial disease modeling and mechanism elucidation to high-throughput compound screening and lead optimization.
Case Study 1: Large-Scale Drug Screening for Sporadic ALS Using iPSC-Derived Motor Neurons
Experimental Background and Rationale
Amyotrophic lateral sclerosis (ALS) presents a formidable challenge for drug development due to its clinical heterogeneity and the predominance (90%) of sporadic cases (SALS) with unknown etiology. Prior to the development of iPSC models, the field relied primarily on models expressing mutations from rare familial forms (FALS), often with nonphysiological expression levels and regulation [83]. This limitation resulted in poor translational success, with more than 160 drugs that showed efficacy in FALS models failing in clinical trials for SALS patients [83]. The establishment of a large-scale iPSC library from 100 SALS patients represented a paradigm shift, enabling population-wide phenotypic screening that captures the biological and genetic diversity of the actual patient population.
Detailed Experimental Methodology
iPSC Library Generation and Quality Control
- Donor Cohort: Skin biopsies were obtained from 100 clinically assessed SALS patients with no family history and 25 healthy controls without neurodegenerative disease history [83].
- Reprogramming Method: Fibroblasts were isolated and reprogrammed using nonintegrating episomal vectors on an automated robotics platform to maximize output uniformity and reproducibility [83].
- Quality Control: All lines underwent rigorous quality assessment including confirmation of genomic integrity, pluripotency markers, and trilineage differentiation potential [83].
- Genetic Characterization: Whole-genome DNA sequencing identified pathogenic variants in known ALS genes in a subset of donors, who were subsequently classified as FALS and excluded from the SALS cohort [83].
Motor Neuron Differentiation and Characterization
Researchers implemented an optimized five-stage spinal motor neuron differentiation protocol adapted from established methods [83]:
- Neural induction using dual SMAD inhibition
- Patterning toward caudal hindbrain and spinal cord fates
- Motor neuron specification with retinoic acid and sonic hedgehog pathway agonists
- Progenitor expansion
- Terminal differentiation and maturation
Characterization Results:
- Purity Assessment: Cultures contained
92.44 ± 1.66%motor neurons (co-expressing ChAT, MNX1/HB9, and Tuj1) [83]. - Contamination Check: Minimal presence of astrocytes (
0.12 ± 0.01%GFAP+ cells) and microglia (0.04 ± 0.02%CD11B+ cells) [83]. - Functional Assessment: Neurons exhibited extensive neurite networks and electrophysiological activity.
Phenotypic Screening and Validation
- Longitudinal Live-Cell Imaging: Cultures were monitored daily using live-cell imaging with a virally delivered nonintegrating motor neuron-specific reporter (HB9-turbo) to track survival and neurite degeneration [83].
- Quantitative Criteria: Highly stringent quantification criteria were implemented to distinguish healthy control from diseased motor neurons [83].
- Donor Correlation: Neurite degeneration phenotypes were correlated with donor survival times to establish clinical relevance [83].
Key Experimental Results and Data Analysis
Table 1: Summary of SALS Motor Neuron Phenotypes in iPSC Model
| Parameter Assessed | Finding in SALS vs. Control | Statistical Significance | Clinical Correlation |
|---|---|---|---|
| Neuronal Survival | Significantly reduced | p < 0.001 | Not specified |
| Neurite Degeneration | Accelerated | p < 0.001 | Correlated with donor survival (p < 0.01) |
| Transcriptional Profile | Significant differential expression | Not specified | Consistent with postmortem ALS spinal cord |
| Pharmacological Response | Riluzole-responsive | p < 0.01 | Consistent with clinical efficacy |
Table 2: Drug Screening Outcomes in SALS iPSC Model
| Compound Category | Number Tested | Number Effective | Success Rate | Key Findings |
|---|---|---|---|---|
| Drugs previously tested in ALS clinical trials | >100 | <5 | <5% | Reflected clinical trial failure rates |
| Effective compounds identified | 3 | 3 | 100% | Riluzole, memantine, baricitinib |
| Combinatorial therapy | 1 combination | 1 effective | 100% | Significantly increased SALS motor neuron survival |
Experimental Workflow Visualization
Research Reagent Solutions
Table 3: Key Research Reagents for iPSC-Derived Motor Neuron Screening
| Reagent / Tool | Specific Example | Function in Experimental Protocol |
|---|---|---|
| Reprogramming System | Non-integrating episomal vectors | Footprint-free somatic cell reprogramming to iPSCs |
| Motor Neuron Differentiation Kit | 5-stage protocol with RA and SAG | Directed differentiation to spinal motor neurons |
| Cell Type Markers | ChAT, MNX1/HB9, Tuj1, GFAP, CD11B | Characterization of purity and cellular composition |
| Viral Reporter | HB9-turbo | Motor neuron-specific labeling for live imaging |
| Live-Cell Imaging System | Longitudinal imaging platform | Continuous monitoring of survival and neurite health |
Case Study 2: Modeling Neurodevelopmental Disorders Using Cortical Neurons
Experimental Background and Rationale
Neurodevelopmental disorders such as autism spectrum disorders (ASD) and schizophrenia (SCZ) present unique challenges for drug discovery due to the complex interplay of genetic risk factors and the inaccessibility of living human neurons for study [103]. iPSC technology enables researchers to capture the complete genetic background of affected individuals, including polygenic risk architectures that cannot be modeled effectively in traditional systems. This case study examines two complementary approaches: village editing for schizophrenia research and dorsal-ventral assembloids for studying interneuron migration.
Detailed Experimental Methodology
Village Editing for Schizophrenia Risk Modeling
- Donor Selection: iPSC lines from 15 donors with low, neutral, or high polygenic risk scores for schizophrenia [4].
- Genetic Manipulation: CRISPR/Cas9 gene editing in a "cell village" format to generate NRXN1 knockouts (KOs) across multiple genetic backgrounds [4].
- Editing Efficiency: Successful generation of heterozygous (33.1%) and homozygous (28.4%) NRXN1 deletions across most donors [4].
- Differentiation Protocol: iPSCs differentiated to cortical excitatory neurons over 28 days with mouse glial support [4].
- Transcriptional Analysis: RNA sequencing to determine NRXN1 KO effects on neuron transcriptomes across different genetic backgrounds [4].
Dorsal-Ventral Assembloid Model for Interneuron Migration
- Model Design: iPSC-derived dorsal and ventral forebrain organoids fused to create assembloids [4].
- Culture Duration: Extended culture period up to 390 days to model postnatal migration events [4].
- Migration Analysis: Time-lapse imaging to track caudal ganglionic eminence (CGE) interneuron migration [4].
- Molecular Characterization: Single-cell spatial transcriptomics and electron microscopy to validate structural architecture [4].
Key Experimental Results and Data Analysis
Table 4: Neurodevelopmental Disorder Modeling Approaches and Outcomes
| Model System | Genetic/Phenotypic Focus | Key Findings | Therapeutic Implications |
|---|---|---|---|
| NRXN1 Village Editing | Schizophrenia polygenic risk | Genetic background deeply influences NRXN1 KO transcriptomic effects | Personalized therapeutic approaches needed for NRXN1-related disorders |
| Cortical-Striatal Assembloids | Interneuron migration defects | Chain migration of interneurons surrounded by astrocytes; requires neuron-astrocyte interactions | New targets for disorders with interneuron deficits (epilepsy, autism) |
| HSAN IV DRG Organoids | NTRK1 mutations in pain insensitivity | Lineage switching between sensory neurons and glial cells; reduced ISLET+/BRN3A+ neurons | Possible strategies to restore sensory neuron differentiation |
Signaling Pathway and Experimental Workflow
Research Reagent Solutions
Table 5: Key Research Reagents for Neurodevelopmental Disease Modeling
| Reagent / Tool | Specific Example | Function in Experimental Protocol |
|---|---|---|
| CRISPR/Cas9 System | NRXN1 targeting guides | Precise gene editing in cell village format |
| Polygenic Risk Score | Schizophrenia PRS calculation | Donor stratification for genetic background effects |
| Neuronal Differentiation Kit | Cortical neuron protocol | Generation of excitatory neurons for transcriptomics |
| Assembloid Protocol | Dorsal-ventral fusion method | Modeling interneuron migration and connectivity |
| Spatial Transcriptomics | Single-cell RNA mapping | Characterization of migration patterns and cell interactions |
Discussion: Integration into Drug Development Pipelines
The case studies presented demonstrate how iPSC-derived neural cells are being systematically integrated across the pharmaceutical development pipeline, from early target identification to preclinical safety assessment. Several key themes emerge from these examples that highlight both the promise and challenges of this technology.
Addressing Clinical Failure Rates Through Human-Relevant Models
The high concordance between clinical trial outcomes and iPSC-based screening results in the SALS study [83] provides compelling evidence for the predictive validity of these models. By accurately reflecting the heterogeneity of patient populations, iPSC-based screening platforms enable more clinically relevant assessment of therapeutic candidates before entering costly clinical trials. Furthermore, the identification of effective drug combinations that work across diverse genetic backgrounds represents a significant advance for complex sporadic disorders [83].
Technical Considerations and Protocol Optimization
The success of iPSC-based drug screening depends critically on protocol standardization and quality control. The SALS study highlighted the importance of:
- Rigorous differentiation protocols yielding >90% pure motor neuron populations [83]
- Longitudinal phenotypic assessment with clinically relevant endpoints [83]
- Large sample sizes to account for donor-to-donor variability [83]
Similarly, the neurodevelopmental studies demonstrated the value of:
- Incorporating polygenic risk背景 rather than focusing exclusively on monogenic mutations [4]
- Extended culture durations to capture later developmental events [4]
- Multi-cellular systems (assembloids) to model cell-cell interactions [4] [30]
Future Directions and Emerging Applications
Recent advances in deterministic reprogramming using technologies like opti-ox enable generation of more consistent, defined human cell types, addressing the batch-to-batch variability that has plagued conventional iPSC differentiation [100]. The integration of artificial intelligence with iPSC-based screening is creating new opportunities for pattern recognition and predictive modeling [104]. Additionally, the development of 3D organoid and assembloid platforms continues to enhance the physiological relevance of these models [30].
iPSC-derived neural cells have evolved from a research tool to a critical component of the pharmaceutical development pipeline. The case studies examined in this whitepaper demonstrate how these models are being used to deconvolute complex disease mechanisms, screen compound libraries with human-relevant readouts, and identify promising therapeutic candidates with increased translational potential. For neurodevelopmental disorders specifically, the ability to model patient-specific genetic backgrounds in developing neural circuits provides unprecedented opportunities for understanding disease mechanisms and developing targeted interventions. As protocols continue to standardize and model complexity increases, iPSC technology is poised to substantially impact the success rate of neuroscience drug development, potentially reversing the historical trend of high failure rates in CNS clinical trials.
The advent of induced pluripotent stem cell (iPSC) technology has revolutionized the study of the human nervous system and its disorders. By enabling the generation of patient-specific neural cells, iPSCs provide an unprecedented platform for modeling neurodevelopmental disorders (NDDs), elucidating disease mechanisms, and conducting drug screening [105] [3]. This technology, which involves reprogramming somatic cells to a pluripotent state using defined factors, allows researchers to capture the unique genetic background of individuals with neurological conditions [105]. The core premise of "disease in a dish" modeling involves generating iPSCs from patients, differentiating them into disease-relevant neural cell types, and comparing their phenotypes to healthy controls to uncover cellular and molecular pathologies [105].
The complexity of iPSC-based models ranges from simple two-dimensional (2D) monocultures to intricate three-dimensional (3D) organoid systems that better mimic the architecture of the human brain [20] [106]. Selecting the appropriate model system and neural cell type is crucial for effectively modeling specific aspects of NDDs. This review provides a comparative analysis of different iPSC-derived neural cell types and model system complexities, focusing on their applications in NDD research. We evaluate the strengths and limitations of each approach, provide detailed methodological insights, and discuss how these models are advancing our understanding of neurodevelopmental disorder mechanisms.
iPSC-Derived Neural Cell Types for Disease Modeling
The in vitro differentiation of iPSCs into various neural cell types leverages principles of developmental biology, utilizing specific morphogens and small molecules to direct cell fate. The following sections detail the generation and application of key neural cell types relevant to NDD research.
Neural Stem Cells (NSCs)
Neural stem cells serve as a multipotent reservoir for generating neurons and glial cells in the brain. iPSC-derived NSCs are typically generated through dual SMAD inhibition using small molecules that target key developmental pathways such as TGFβ and BMP [105]. Common compounds used include SB431542 (a TGFβ inhibitor), dorsomorphin (a BMP inhibitor), and CHIR99021 (a GSK3 inhibitor that activates WNT signaling) [105]. These molecules work synergistically to efficiently induce a neural fate in pluripotent cells within approximately one week, resulting in cells that express characteristic NSC markers like N-Cadherin, SOX2, and NESTIN [105].
iPSC-derived NSCs have been particularly valuable for studying the early developmental aspects of NDDs. For instance, NSCs derived from patients with Parkinson's disease carrying LRRK2 mutations exhibited nuclear architecture defects and increased proteasomal stress, revealing novel disease phenotypes [105]. In studies of neuropsychiatric disorders such as schizophrenia and autism associated with 15q11.2 copy number variations, patient-derived NSCs showed deficits in cytoskeleton organization mediated by CYFIP1, providing insights into how genetic lesions disrupt fundamental cellular processes during development [105]. Similarly, NSCs from individuals with William syndrome demonstrated proliferation and survival defects that were traced to abnormalities in the FZD9 gene [105].
A significant consideration when using NSCs to model age-related disorders is their inherently fetal character. To address this limitation, researchers have employed progerin, MG132, and concanamycin A to induce aging-related phenotypes in iPSC-derived neural cells, thereby enhancing the relevance of these models for late-onset conditions [105].
Neurons
The generation of specific neuronal subtypes from iPSCs is achieved by recapitulating regional patterning signals that occur during embryonic development. These protocols typically involve the sequential application of morphogens that pattern the neuroepithelium along its rostro-caudal and dorso-ventral axes [105].
Table 1: Regional Patterning of iPSC-Derived Neurons
| Neuronal Subtype | Key Patterning Factors | Associated Disorders |
|---|---|---|
| Glutamatergic Neurons | Default differentiation in absence of exogenous morphogens; endogenous WNTs [105] | Schizophrenia, Autism Spectrum Disorders [107] |
| GABAergic Neurons | SHH activation + WNT inhibition [105] | Schizophrenia, Epilepsy [107] |
| Basal Forebrain Cholinergic Neurons (BFCNs) | SHH/Purmorphamine + Nerve Growth Factor (NGF) [105] | Alzheimer's Disease, Cognitive Disorders |
| Spinal Motor Neurons (MNs) | SHH (ventralization) + Retinoic Acid (caudalization) [105] [108] | Amyotrophic Lateral Sclerosis, Spinal Muscular Atrophy [83] |
The functionality of iPSC-derived neurons is typically validated through multiple methods, including immunostaining for subtype-specific markers, patch-clamp electrophysiology to demonstrate action potential generation, and calcium imaging to assess network activity [108]. For example, motor neurons derived using optimized protocols express markers such as ChAT, MNX1/HB9, and β-tubulin III, with functional maturation evident by approximately 7-8 weeks in culture [83].
These neuronal models have been instrumental in revealing disease-specific phenotypes. In studies of amyotrophic lateral sclerosis (ALS), patient-derived motor neurons show reduced survival and accelerated neurite degeneration, correlating with donor survival time [83]. For autism spectrum disorder and schizophrenia, patient-derived neurons have demonstrated alterations in neurite outgrowth, synaptic connectivity, and transcriptional profiles that distinguish these conditions from each other and from healthy controls [107].
Glial Cells
Glial cells, including astrocytes and oligodendrocytes, play crucial roles in neural development and function, and their dysfunction has been implicated in various NDDs. The differentiation of iPSCs into glial cells generally follows a longer timeline than neuronal differentiation, with astrocyte progenitors appearing after approximately 4 weeks and mature GFAP-expressing astrocytes emerging around 3 months [109]. Oligodendrocytes typically begin to appear after 4 months of differentiation [109].
iPSC-derived glia have been used to model cell-type-specific contributions to NDDs. In Fragile X syndrome, for example, astrocyte generation has been shown to vary among iPSC lines from different patients, suggesting that glial dysfunction may contribute to the disorder's pathophysiology [20]. Similarly, in Alexander disease, a leukodystrophy characterized by astrocyte dysfunction, patient-derived glial models have provided insights into disease mechanisms [20].
Model System Complexities: From 2D Cultures to 3D Organoids
The complexity of iPSC-based neural models has evolved significantly, from simple 2D monocultures to sophisticated 3D organoid systems that better recapitulate tissue architecture and cell-cell interactions.
2D Monocultures
Two-dimensional monocultures represent the most reductionist approach to modeling neural cells in vitro. These systems typically involve differentiating iPSCs into a specific neural cell type (e.g., neurons, astrocytes) and maintaining them as a monolayer on coated surfaces [109]. The primary advantages of 2D systems include their simplicity, reproducibility, and suitability for high-throughput screening applications. For instance, large-scale drug screening in iPSC-derived motor neurons from 100 sporadic ALS patients was conducted using 2D cultures, identifying potential combinatorial therapies [83].
However, 2D systems have significant limitations. They lack the complex cellular interactions and tissue architecture of the developing brain, and may not fully recapitulate the maturation states observed in vivo. Additionally, neural differentiation efficiency in 2D cultures can be variable across iPSC lines, with some lines showing poor response to neural inducers regardless of the reprogramming method used [109].
Co-culture Systems
Co-culture systems involve growing two or more neural cell types together to study their interactions. These systems provide a more physiologically relevant context than monocultures, as they allow for the investigation of cell-cell signaling and non-cell-autonomous disease mechanisms. For example, co-culturing neurons with astrocytes has revealed important roles for glial cells in modulating synaptic function and neuronal survival in various NDD models [20].
3D Brain Organoids and Assembloids
Three-dimensional brain organoids represent a significant advancement in complexity, as they self-organize to recapitulate aspects of the developing human brain's architecture and cellular diversity [106]. These models are generated by embedding iPSC aggregates in extracellular matrix substitutes and exposing them to patterning factors that promote regional identity. Recent protocols have improved the reproducibility and maturity of organoids, addressing earlier limitations such as necrosis and insufficient neuronal maturation [106].
Assembloids represent a further advancement, involving the fusion of region-specific organoids to model interactions between different brain areas [106]. For instance, cortical organoids fused with thalamic organoids have been used to reconstruct corticothalamic circuitry, revealing insights into the neural connectivity underlying neurodevelopmental processes [106].
Table 2: Comparison of iPSC-Based Neural Model Systems
| Model System | Key Features | Advantages | Limitations | Applications in NDDs |
|---|---|---|---|---|
| 2D Monocultures | Single cell type; planar growth [109] | High reproducibility; suitable for high-throughput screening; ease of manipulation and analysis [83] | Limited cellular interactions; lacks tissue architecture; may not fully mature [109] | Drug screening (e.g., ALS [83]); mechanistic studies of cell-autonomous defects |
| Co-culture Systems | Multiple cell types in 2D [20] | Enables study of cell-cell interactions; more physiologically relevant than monocultures | Still lacks 3D architecture; challenging to control cell ratios | Modeling non-cell-autonomous effects (e.g., neuron-astrocyte interactions in FXS [20]) |
| 3D Organoids | Multiple cell types in 3D; self-organization [106] | Recapitulates some tissue architecture; contains diverse cell types; better maturation potential | Heterogeneity between organoids; core necrosis issues; complex analysis | Modeling cortical development; studying disorders of brain organization [20] |
| Assembloids | Fused region-specific organoids [106] | Models circuit formation between brain regions; enables studying long-range projections | Technically challenging; variability in fusion efficiency | Studying neural connectivity defects in NDDs; modeling corticostriatal circuits [106] |
The following workflow diagram illustrates the general process for generating different iPSC-derived neural models:
Experimental Protocols for iPSC Neural Differentiation
Neural Stem Cell Differentiation Protocol
The following protocol efficiently differentiates iPSCs into NSCs using small molecule inhibition [105]:
- Initial Seeding: Plate iPSCs as single cells on Matrigel-coated plates in mTeSR or Essential 8 medium with 10µM ROCK inhibitor.
- Dual SMAD Inhibition: Once cells reach 70-80% confluence, switch to neural induction medium containing:
- 1µM Dorsomorphin (BMP inhibitor)
- 10µM SB431542 (TGFβ inhibitor)
- 3µM CHIR99021 (GSK3β inhibitor/WNT activator)
- 0.5µM Compound E (Notch inhibitor)
- Culture Duration: Maintain cells in induction medium for 7-10 days with daily medium changes.
- NSC Expansion: Passage cells using Accutase and plate on poly-ornithine/laminin-coated plates in NSC expansion medium containing LIF to maintain pluripotency.
The resulting NSCs should express markers including PAX6, SOX1, N-Cadherin, and NESTIN, and possess the capacity to differentiate into multiple neural lineages.
Motor Neuron Differentiation Protocol
This rapid protocol generates functional spinal motor neurons suitable for neural injury modeling and disease studies [108]:
- Neural Induction: Form embryoid bodies (EBs) in low-adhesion plates for 7 days in neural induction medium.
- Patterning: At day 7, plate EBs on poly-ornithine/laminin-coated plates in motor neuron patterning medium containing:
- 1µM Retinoic Acid (RA) for caudalization
- 1µM Purmorphamine (SHH agonist) for ventralization
- Maturation: From day 14 onwards, switch to motor neuron maturation medium containing:
- BDNF, GDNF, and CNTF (10-20ng/mL each)
- Ascorbic acid (200µM)
- cAMP (1µM)
- Functional Validation: By day 28-35, neurons should exhibit spontaneous electrical activity, express markers HB9, ISL1, and ChAT, and demonstrate typical electrophysiological properties.
The following diagram illustrates the key signaling pathways manipulated during neural differentiation:
The Scientist's Toolkit: Essential Research Reagents
Successful differentiation and analysis of iPSC-derived neural models require carefully selected reagents and tools. The following table details essential components for iPSC-based neural differentiation and phenotyping:
Table 3: Essential Research Reagents for iPSC Neural Differentiation and Analysis
| Reagent Category | Specific Examples | Function/Purpose |
|---|---|---|
| Reprogramming Factors | OCT4, SOX2, KLF4, MYC (OSKM) [3] | Conversion of somatic cells to pluripotent state; foundational for iPSC generation |
| Neural Induction Compounds | SB431542 (TGFβ inhibitor), Dorsomorphin (BMP inhibitor) [105] | Dual SMAD inhibition for efficient neural induction from pluripotent state |
| Patterning Molecules | CHIR99021 (GSK3 inhibitor/WNT activator), Retinoic Acid, Purmorphamine (SHH agonist) [105] [108] | Regional specification of neural progenitors (rostral/caudal, dorsal/ventral) |
| Growth Factors | BDNF, GDNF, CNTF, NGF [105] [108] | Support neuronal survival, maturation, and maintenance in culture |
| Cell Surface Markers | PAX6, SOX1, NESTIN (NSCs); βIII-tubulin, MAP2 (neurons); GFAP (astrocytes); O4 (oligodendrocytes) [109] | Identification and purification of specific neural cell types |
| Functional Assay Tools | Calcium indicators (e.g., Fluo-4), Patch clamp reagents, Multi-electrode arrays (MEAs) [108] | Assessment of electrophysiological properties and network activity |
| Gene Editing Tools | CRISPR/Cas9, TALENs, ZFNs [20] | Generation of isogenic controls; introduction or correction of disease mutations |
Applications in Neurodevelopmental Disorder Research
iPSC-derived neural models have provided unprecedented insights into the pathophysiology of NDDs. The following sections highlight key applications across different model complexities.
Disease Mechanism Elucidation
iPSC models have been particularly valuable for understanding the cellular and molecular mechanisms underlying NDDs. In Fragile X syndrome, iPSC-derived neurons with hypermethylated FMR1 promoters showed aberrant differentiation and defective neurite outgrowth, which could be rescued by demethylation of CGG repeats using CRISPR/Cas9 genome editing [20]. In schizophrenia models, transcriptomic analyses of patient-derived neurons have identified dysregulation of genes involved in WNT signaling (LEF1) and neurodevelopmental pathways (ERBB3) [107].
For ALS, large-scale studies using iPSC-derived motor neurons from 100 sporadic patients have confirmed reduced neuronal survival and accelerated neurite degeneration, validating the relevance of these models for the predominant sporadic form of the disease [83]. These models have also recapitulated TDP-43 pathology and transcriptional profiles consistent with postmortem spinal cord tissues from ALS patients [83].
Drug Screening and Therapeutic Development
The pharmaceutical applications of iPSC-derived neural models are extensive, particularly for high-throughput compound screening. In the ALS field, screening of drugs previously tested in clinical trials revealed that 97% failed to mitigate neurodegeneration in patient-derived motor neurons, reflecting the high failure rate of ALS clinical trials and validating the predictive value of these models [83]. This approach identified a promising therapeutic combination of baricitinib, memantine, and riluzole that significantly increased motor neuron survival across diverse SALS donors [83].
For neuropsychiatric disorders like schizophrenia and autism, iPSC-derived neuronal models have been used to identify compounds that reverse disease-associated phenotypes, such as altered neuronal connectivity and transcriptional dysregulation [107]. The ability to test drug responses in patient-specific cells also opens possibilities for personalized medicine approaches in NDDs.
Assessment of Selective Neuronal Vulnerability
Single-cell RNA sequencing technologies applied to iPSC-derived neural models have provided insights into the molecular basis of selective neuronal vulnerability in NDDs [110]. By profiling individual cells within heterogeneous cultures or organoids, researchers can identify cell-type-specific responses to genetic risk factors and environmental stressors. This approach has revealed how specific neuronal subtypes are disproportionately affected in conditions like autism, schizophrenia, and neurodegenerative disorders [110].
The systematic comparison of iPSC-derived neural cell types and model system complexities presented in this review highlights the remarkable progress in modeling neurodevelopmental disorders. From simple 2D monocultures to complex 3D assembloids, each model system offers unique advantages and limitations for addressing specific research questions. The choice of model should be guided by the particular aspect of the disorder being investigated, with consideration of the trade-offs between physiological relevance, reproducibility, and scalability.
As iPSC technologies continue to advance, we can expect further refinement of differentiation protocols, improved maturation of neural cells, and increased complexity of 3D model systems. The integration of single-cell multi-omics, high-content imaging, and functional analyses will provide increasingly comprehensive insights into disease mechanisms. For the field of neurodevelopmental disorder research, these developments promise to accelerate the identification of novel therapeutic targets and the development of effective interventions for these complex conditions.
Conclusion
iPSC technology has fundamentally transformed our approach to studying neurodevelopmental disorders, providing unprecedented access to living human neural cells with disease-relevant genetic backgrounds. The evolution from simple 2D cultures to sophisticated 3D organoids and multi-organ systems has enabled more accurate recapitulation of complex disease phenotypes and developmental processes. While challenges remain in standardization, maturation, and scalability, emerging technologies in gene editing, automation, and machine learning are rapidly addressing these limitations. The successful application of iPSC-derived neural models in drug discovery and the establishment of regulatory pathways for their use signal a promising future where these human-centric systems will accelerate the development of targeted therapies for neurodevelopmental disorders, ultimately bridging the long-standing translational gap between preclinical research and clinical benefit for patients.
References
- 1. Animal Disease Models and Patient-iPS-Cell-Derived In ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10045735/]
- 2. Modeling Brain Disorders Using Induced Pluripotent Stem Cells [https://pmc.ncbi.nlm.nih.gov/articles/PMC7263088/]
- 3. Induced pluripotent stem cells (iPSCs) [https://www.nature.com/articles/s41392-024-01809-0]
- 4. Modeling Human Neural Development and Disease with iPSCs — International Society for Stem Cell Research [https://www.isscr.org/upcoming-programs/poster-award-webinar]
- 5. Novel human iPSC models of neuroinflammation in neurodegenerative disease and regenerative medicine - PubMed [https://pubmed.ncbi.nlm.nih.gov/39307583/]
- 6. Qualitative and Quantitative Comparison of the Proteome of ... [https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0100874]
- 7. Frontiers | Editorial: Advancing Neurodevelopmental Disorder Models with Human iPSC and Multi-Omics Integration [https://www.frontiersin.org/journals/neuroscience/articles/10.3389/fnins.2025.1724360/abstract]
- 8. Somatic Cell Reprogramming for Nervous System Diseases [https://pmc.ncbi.nlm.nih.gov/articles/PMC10046178/]
- 9. Direct Lineage Reprogramming: Strategies, Mechanisms ... [https://www.sciencedirect.com/science/article/pii/S1934590915000144]
- 10. Quantitative Analysis of Human Pluripotency and Neural ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5032292/]
- 11. Analysis of neuronal activity impairment in iPSC-derived ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12546973/]
- 12. iPSC-modelling reveals genetic associations and ... [https://www.nature.com/articles/s41398-025-03509-x]
- 13. Induced Pluripotent Stem Cells (iPSC) and Their Use in ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12590075/]
- 14. Induced pluripotency: history, mechanisms, and applications [https://pmc.ncbi.nlm.nih.gov/articles/PMC2956203/]
- 15. A comprehensive analysis of induced pluripotent stem cell ... [https://www.frontiersin.org/journals/cell-and-developmental-biology/articles/10.3389/fcell.2025.1593207/full]
- 16. Overcoming Scale-Up Challenges in iPSC Production [https://www.carrbiosystems.com/resources/publications/post/overcoming-scale-up-challenges-in-ipsc-production]
- 17. Patient-Derived Induced Pluripotent Stem Cells (iPSCs) and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7922555/]
- 18. Editorial: Advancing neurodevelopmental disorder models ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12647108/]
- 19. The Use of Induced Pluripotent Stem Cell Technology to ... [https://www.sciencedirect.com/science/article/pii/S187874792301601X]
- 20. Patient-derived iPSC modeling of rare neurodevelopmental ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7962756/]
- 21. Visualization of morphological categories of colonies for ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6134894/]
- 22. 3D and organoid culture in research: physiology ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8973959/]
- 23. Current Development of iPSC-Based Modeling in ... [https://www.mdpi.com/1422-0067/26/8/3774]
- 24. Generation of 3D Human iPSC-Derived Multi-Cell Type ... [https://pubmed.ncbi.nlm.nih.gov/41230551/]
- 25. A Study Comparing 2D Monolayers with 3D Organoid ... [https://pubmed.ncbi.nlm.nih.gov/33513360/]
- 26. Emerging brain organoids: 3D models to decipher, identify ... [https://www.sciencedirect.com/science/article/pii/S2452199X25000258]
- 27. Induced Pluripotent Stem Cell (iPSC) Industry Trends for ... [https://bioinformant.com/ipsc-industry-trends/]
- 28. A high efficiency precision genome editing method with ... [https://www.nature.com/articles/s41598-024-60766-4]
- 29. The hidden risks of CRISPR/Cas: structural variations and ... [https://www.nature.com/articles/s41467-025-62606-z]
- 30. Brain organoids: a new paradigm for studying human ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12620469/]
- 31. CMN Weekly (26 September 2025) [https://crisprmedicinenews.com/news/cmn-weekly-26-september-2025-your-weekly-crispr-medicine-news/]
- 32. CRISPR Clinical Trials: A 2025 Update [https://innovativegenomics.org/news/crispr-clinical-trials-2025/]
- 33. World's First Patient Treated with Personalized CRISPR ... [https://www.chop.edu/news/worlds-first-patient-treated-personalized-crispr-gene-editing-therapy-childrens-hospital]
- 34. Patch-clamp and multi-electrode array electrophysiological ... [https://pubmed.ncbi.nlm.nih.gov/33899023/]
- 35. Multielectrode Array [https://www.axionbiosystems.com/multielectrode-array]
- 36. Using Induced Pluripotent Stem Cell-Derived Neurons to ... [https://www.frontiersin.org/journals/neurology/articles/10.3389/fneur.2018.00237/full]
- 37. Intracellular Calcium Measurements for Functional ... [https://pubmed.ncbi.nlm.nih.gov/26126448/]
- 38. Functional network properties derived from wide-field ... [https://www.sciencedirect.com/science/article/pii/S1053811922008564]
- 39. Simultaneous multi-patch-clamp and extracellular-array ... [https://www.nature.com/articles/srep36228]
- 40. Neuronal impact of patient-specific aberrant NRXN1α ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7451045/]
- 41. Schizophrenia-associated NRXN1 deletions induce ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10290702/]
- 42. Article Human Neuropsychiatric Disease Modeling using ... [https://www.sciencedirect.com/science/article/pii/S1934590915003161]
- 43. NRXN1-related severe neurodevelopmental disorder ... [https://www.orpha.net/en/disease/detail/600663]
- 44. A Comprehensive Functional Analysis of NTRK1 Missense ... [https://pubmed.ncbi.nlm.nih.gov/27676246/]
- 45. A guide to the organ-on-a-chip [https://www.nature.com/articles/s43586-022-00118-6]
- 46. Organ-on-a-Chip systems for new drugs development - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC8920106/]
- 47. Human organs-on-chips for disease modelling, drug ... [https://www.nature.com/articles/s41576-022-00466-9]
- 48. Developer's Guide to an Organ-on-Chip Model - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC9667877/]
- 49. Using Organ-on-a-Chip and iPSC Technology to Model ... [https://emulatebio.com/event/using-organ-on-a-chip-and-ipsc-technology-to-model-neurological-diseases/]
- 50. Organoids-on-Chips Technology: Unveiling New ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12072464/]
- 51. A Possible Reservoir of Young Cells to Prevent Brain Aging ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6571946/]
- 52. Hyper-maturity and accelerated aging in the hippocampus ... [https://www.nature.com/articles/s41386-025-02237-6]
- 53. Mechanisms underlying neurodevelopmental disorders [https://www.sciencedirect.com/science/article/abs/pii/S0959438825000741]
- 54. Scientists uncover excessive maturation and aging in the ... [https://www.news-medical.net/news/20251027/Scientists-uncover-excessive-maturation-and-aging-in-the-hippocampus.aspx]
- 55. hydroxynorketamine and reelin: an exploratory study [https://www.nature.com/articles/s41398-025-03724-6]
- 56. Neurogenesis in aging and age-related neurodegenerative ... [https://www.sciencedirect.com/science/article/abs/pii/S1568163722000782]
- 57. Towards a quality control framework for cerebral cortical ... [https://www.nature.com/articles/s41598-025-14425-x]
- 58. Validating human induced pluripotent stem cell-specific ... [https://www.sciencedirect.com/science/article/pii/S1465324924006108]
- 59. A Protocol for Culture and Characterization of Human ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10506163/]
- 60. Optimization of Transcription Factor-Driven Neuronal ... [https://link.springer.com/article/10.1007/s12015-025-10845-4]
- 61. iPSCs and iPSC-derived cells as a model of human ... [https://www.nature.com/articles/s41467-025-56569-4]
- 62. Modeling developmental neuropsychiatric disorders with iPSC ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4738093/]
- 63. Opportunities and limitations for studying neuropsychiatric ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10213114/]
- 64. Bioprocessing considerations for generation of iPSCs ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12288090/]
- 65. Radical Bioprocessing Efficiencies and Cost Reductions [https://www.pharmasalmanac.com/articles/radical-bioprocessing-efficiencies-and-cost-reductions-the-next-wave-of-biopharmaceutical-innovation]
- 66. Neurodevelopmental disorders—high-resolution rethinking ... [https://www.nature.com/articles/s41380-022-01876-1]
- 67. Reduce Staffing Costs and Boost Access with Cell Therapy ... [https://www.genengnews.com/topics/bioprocessing/reduce-staffing-costs-and-boost-access-with-cell-therapy-automation/]
- 68. The Impact of Intelligent Automation on Cost Savings [https://integranxt.com/blog/impact-of-intelligent-automation-on-cost-savings/]
- 69. Reducing Development Costs and Timelines in ... [https://www.idbs.com/knowledge-base/reducing-development-costs-and-timelines-in-bioprocessing/]
- 70. Automated high-content imaging in iPSC-derived neuronal ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10602900/]
- 71. Top 10 Machine Learning Frameworks to Use In 2025 [https://www.igmguru.com/blog/machine-learning-frameworks]
- 72. Top 10 Machine Learning Frameworks in 2025 [https://www.geeksforgeeks.org/blogs/machine-learning-frameworks/]
- 73. A Novel Automated High-Content Analysis Workflow ... [https://www.sciencedirect.com/science/article/pii/S2472555222071519]
- 74. iPhemap: an atlas of phenotype to genotype relationships ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5731211/]
- 75. TableGPT2: A Large Multimodal Model with Tabular Data ... [https://arxiv.org/html/2411.02059v1]
- 76. Genetic and epigenetic variations in iPSCs: potential ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3760008/]
- 77. Their Role in iPSC Reprogramming and Differentiation [https://www.reprocell.com/blog/epigenetic-modifications-their-role-in-ipsc-reprogramming-and-differentiation]
- 78. Section 1: Basic Characterization [https://www.isscr.org/basic-research-standards/basic-characterization]
- 79. Cell Banking Best Practices [https://www.wicell.org/blog/cell-banking-best-practices/]
- 80. Statistical Methods for Multi-Omics Analysis in ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12562612/]
- 81. Cell-specific expression biases in human cortex of genes ... [https://www.nature.com/articles/s41598-025-05117-7]
- 82. Postsynaptic structure formation of human iPS cell-derived ... [https://molecularbrain.biomedcentral.com/articles/10.1186/s13041-021-00851-1]
- 83. Large-scale drug screening in iPSC-derived motor neurons ... [https://www.nature.com/articles/s41593-025-02118-7]
- 84. Human Pluripotent Stem Cell-Derived Models and Drug ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8193821/]
- 85. Induced Pluripotent Stem Cells in Drug Discovery and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10889263/]
- 86. Induced pluripotent stem cells: applications in regenerative ... [https://www.frontiersin.org/journals/cell-and-developmental-biology/articles/10.3389/fcell.2015.00002/full]
- 87. Screening for variable drug responses using human iPSC ... [https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0323953]
- 88. Multi-lineage Human iPSC-Derived Platforms for Disease ... [https://www.sciencedirect.com/science/article/pii/S1934590920300643]
- 89. P18-119 A 3D Human iPSC-Derived Myelination Model for ... [https://www.sciencedirect.com/science/article/pii/S0378427425021484]
- 90. Pluripotent Stem Cells in Developmental Toxicity Testing [https://pmc.ncbi.nlm.nih.gov/articles/PMC6111785/]
- 91. FDA Modernization Act 2.0: transitioning beyond animal ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10617761/]
- 92. An Overview of the FDA Modernization Act 2.0 [https://www.mattek.com/news-resources/overview-of-fda-modernization-act/]
- 93. Human induced pluripotent stem cells for modeling ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5782822/]
- 94. Patient-derived iPSC modeling of rare ... [https://www.sciencedirect.com/science/article/abs/pii/S0149763420307004]
- 95. Great Expectations: Induced pluripotent stem cell ... [https://www.medsci.org/v18p0459.htm]
- 96. FDA Accelerates Approvals to Reduce Animal Testing [https://www.mwe.com/insights/quit-monkeying-around-fda-offers-faster-approval-times-in-bid-to-phase-out-animal-testing/]
- 97. FDA Announces Plan to Phase Out Animal Testing ... [https://www.fda.gov/news-events/press-announcements/fda-announces-plan-phase-out-animal-testing-requirement-monoclonal-antibodies-and-other-drugs]
- 98. FDA Announces Plan to Phase Out Animal Testing ... [https://barragan.house.gov/2025/04/16/jdsupra-fda-announces-plan-to-phase-out-animal-testing-requirement/]
- 99. FDA Announces Plan to Phase Out Animal Testing ... [https://www.hklaw.com/en/insights/publications/2025/04/fda-announces-plan-to-phase-out-animal-testing-requirement]
- 100. Cell-based drug discovery: Why human iPSC-derived ... [https://www.bit.bio/blog/cell-based-drug-discovery-with-human-ipsc-models]
- 101. Utility of iPSC-Derived Cells for Disease Modeling, Drug ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9180434/]
- 102. How iPS cells are changing drug development and ... [https://www.biopharma-excellence.com/2020-2-28-how-ips-cells-are-changing-drug-development-and-disease-modelling/]
- 103. Modeling neurodevelopmental disorders using human ... [https://www.sciencedirect.com/science/article/abs/pii/S0959438812000621]
- 104. Drug discovery research with the iPSC models of ... [https://www.sciencedirect.com/science/article/pii/S0168010225001683]
- 105. Modeling neurological diseases using iPSC-derived neural cells: iPSC modeling of neurological diseases - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC6029980/]
- 106. building higher-order complexity and neural circuitry models [https://www.sciencedirect.com/science/article/pii/S0167779925000460]
- 107. The potential of induced pluripotent stem cells for ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7780807/]
- 108. Rapid and efficient differentiation of functional motor ... [https://www.sciencedirect.com/science/article/pii/S1873506118302289]
- 109. Neural differentiation of human induced pluripotent stem cells ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC2840097/]
- 110. Single-cell and spatial transcriptomics: deciphering brain ... [https://www.nature.com/articles/s41582-023-00809-y]